
lncRNA NEAT1通过靶向抑制miR-29a促进低氧低糖诱导的肺纤维化进程的机制研究
2021-04-08 08:29段效军李林瑞陈艳萍
实用药物与临床 2021年3期
张 喜,段效军,李林瑞,陈艳萍
0 引言
肺纤维化是一种常见的由慢性阻塞性肺疾病、严重感染、肺损伤、环境因素等引起的肺疾病[1]。缺氧缺血在许多生物学或者病理学过程中起着至关重要的作用[2]。当肺处于缺氧缺血状态时,肺泡上皮细胞由于应激发生肺纤维化,最终可能造成癌变[3-4]。目前,临床上还没有专门用于预防缺血缺氧导致肺纤维化的药物,治疗策略也仅限于支持性治疗。因此,从缺血缺氧引发的纤维化的一系列因子作用环节中寻找有效的治疗靶点,为预防和改善缺血缺氧介导的肺纤维化具有重大意义。
lncRNA一般通过对miRNA的sponge作用调控目标蛋白,进而影响细胞的生物学改变[5]。近年来,lncRNA在纤维化中的作用受到广泛关注,但在调控肺纤维化的研究报道较少,且主要集中于博来霉素诱导的肺纤维化[6]。在低氧诱导的肺纤维化发生过程中,lncRNAs是否也发挥重要的调控作用,还有待深入研究。NEAT1是新近被发现的位于亚核体结构“paraspeckles”周围的lncRNA[7]。研究发现,lncRNA NEAT1可通过调控miR-122和KLF6加速肝纤维化的进程[8]。此外,lncRNA NEAT1可通过激活Akt/mTOR信号通路加速糖尿病肾病的增殖和纤维化[9]。研究者进行了与肺纤维化发病机制相关的研究,旨在确定治疗靶点,但其分子机制仍不明确。因此,了解lncRNA NEAT1在低氧诱导的肺纤维化进展中的作用,可以为肺纤维化的治疗提供新的靶点。
MicroRNAs (miRNAs)是一类以mRNA为靶点并调节翻译抑制的大型非编码RNA分子[10]。miRNAs参与许多生物学过程,如发育时间[11]、细胞增殖[12]、凋亡[13]、神经元模式[14]、造血和器官发育[15]等。近年研究显示,miR-29家族在纤维化中起关键作用,miR-29a可通过抑制CD4+T细胞的分化阻止肝纤维化的进展[16-17]。然而,miR-29a在肺纤维化过程中的变化尚无报道。因此,系统、全面地描述lncRNA NEAT1和miR-29a之间的相互作用和调控模式,可以增强我们对肺纤维化发病机制的理解。
本研究旨在研究lncRNA NEAT1和miR-29a在低氧低糖诱导的肺纤维化中的表达以及lncRNA NEAT1是否可以调控miR-29a的表达,从而影响低氧诱导的肺纤维化进程,并阐明其潜在的调控机制。
1 材料与方法
1.1 材料
1.1.1 临床样本 收集湖南省儿童医院健康者和缺血再灌注引起急性肺损伤患者的血清样本各20份。所有患者均签署了知情同意书,所有实验均获得医院伦理委员会的批准与监督。每份血清样本都保存在-80 ℃直至使用。
1.1.2 动物 40只8周龄C57/B6野生型小鼠由湖南省实验动物中心提供。所有动物实验均按照国家实验动物管理法规和动物实验伦理委员会规定进行。
1.1.3 细胞 人肺泡II型上皮细胞A549和HEK-293T细胞购自中国科学院上海细胞生物学研究所。
1.1.4 试剂 DMEM和RPMI 1640培养液和胎牛血清(FBS)购自美国Thermo Fisher Scientific公司;TRIzol试剂来自美国Invitrogen公司;SYBR Premix EX TaqTMI试剂盒购自日本TaKaRa公司;抗col1、col3a1、α-SMA和TGF-β抗体来自美国Cell Signaling Technologies公司;多聚甲醛、HE染料和Masson染料购自北京Solarbio公司;裂解液等生化试剂购自上海罗氏公司;慢病毒介导的sh-NEAT1、anti-miR-29a以及阴性对照(NC)载体购自中国上海Genechem公司;羟脯氨酸试剂盒和CCK-8试剂盒购自南京建城生物工程研究所;双荧光素酶报告试剂盒购自美国Promega公司。
1.2 方法
1.2.1 细胞的培养、模型构建及转染 细胞的培养:A549细胞用含10%胎牛血清的RPMI 1640培养基培养,HEK-293T细胞用含10%胎牛血清的DMEM培养基培养,都置于37 ℃、5%CO2的细胞培养箱中培养,48 h全换液1次,保证细胞正常生长所需的营养,待细胞密度长至90%,使用0.25%胰酶消化按1∶3进行传代。选取对数生长期的细胞进行后续实验。
肺纤维化细胞模型的构建:将A549细胞置于RPMI 1640+10%胎牛血清的培养液中,放入37 ℃、5%CO2的培养箱内培养,作为对照组,细胞置于无糖平衡盐溶液(pH=7.4)培养,放入37 ℃、5%CO2、1%O2的缺氧培养箱内培养作为模型组。同时在模型组的细胞中转染sh-NC和sh-NEAT1 病毒作为模型+sh-NC组和模型+sh-NEAT1组。
细胞转染:将肺泡上皮细胞(A549)接种在培养皿中,培养18~24 h,待细胞密度至2×105/孔时,用含有6 μg/ml polybrene的新鲜培养液替换旧培养液,进行慢病毒转染,37 ℃培养24 h后进行全换液,继续培养24 h后进行后续实验。
1.2.2 Hoechst染色 使用Hoechst染色试剂盒,按照制造商的说明书对A549细胞进行细胞凋亡检测。取洁净盖玻片在70%乙醇中浸泡5 min,用PBS洗涤3次,再用细胞培养液洗涤1次,将盖玻片置于6孔板,接种细胞培养过夜。对细胞进行转染处理后,吸尽培养液,加入Hoechst固定液0.5 ml,固定10 min,吸走固定液,用PBS洗2次,3 min/次,加入Hoechst染色液0.5 ml,孵育5 min,弃染色液,用PBS洗2次,3 min/次,滴1滴防淬灭封片剂于载玻片上,盖上有细胞的盖玻片。在荧光显微镜下进行分析。
1.2.3 Western blot 用RIPA裂解液裂解肺泡上皮细胞和肺组织,提取蛋白质并进行蛋白定量和煮样。配置10%的SDS-PADE琼脂糖凝胶进行电泳(80 V 20 min,100 V 1 h),电泳结束后进行电转(100 V 150 min),再用5%的牛奶对膜进行室温封闭1 h,用1×TBST洗3次,10 min/次,孵育一抗4 ℃过夜,次日用1×TBST洗3次膜,10 min/次,再室温孵育二抗1 h,用1×TBST洗3次膜,10 min/次,进行显影。最后使用Image J软件对Western blot条带进行定量,并进行归一化。主要一抗有β-actin(抗兔)、col1(抗兔,1∶1 000)、col3a1(抗兔,1∶1 000)、α-SMA(抗兔,1∶1 000)和TGF-β(抗兔,1∶1 000),二抗是山羊抗兔IgG抗体(1∶1 000)。
1.2.4 实时定量PCR(qRT-PCR) 用TRIzol法提取细胞或组织的总RNA。细胞去除培养基,每皿直接加入1 ml TRIzol,反复吹打转移至RNA-free的EP管,室温静置5 min,每管加入250 μl氯仿,室温静置3 min,14 000 g,4 ℃离心15 min。样品分为3层,水相是在室温下与500 μl异丙醇混合10 min。离心机在14 000 g,4 ℃ 离心10 min,取上层清液加入1 ml乙醇洗RNA,在10 000 g,4 ℃离心5 min,弃上清加入10 μl RNA-free水。用NanoDrop 8000测定提取的RNA的浓度和纯度。再反转为cDNA,使用SYBR Premix EX TaqTM试剂盒进行RT-PCR检测mRNA的相对表达水平。采用2-△△Ct法计算相对表达水平。引物序列见表1。

表1 引物序列
1.2.5 缺血缺氧肺损伤动物模型的构建 动物随机分为对照组、模型组、模型+sh-NC组、模型+sh-NEAT1组。实验前大鼠禁食不禁水,戊巴比妥钠腹腔注射麻醉,静脉留置导管,气管插管,接动物呼吸机。模型组剪开颈部皮肤,经右颈静脉注射肝素。平卧位经左五肋间进胸,暴露左肺门,以无损伤血管夹夹闭左肺门。对照组开胸后仅游离肺门,不做夹闭。临时关闭胸腔,后开放血管夹并逐层关胸,再灌注6 h。模型组+sh-NC组和模型组+sh-NEAT1组的小鼠再经尾静脉注射接受病毒sh-NC和sh-NEAT1溶液,病毒处理35 d后对老鼠进行安乐死,经肺循环灌流后肺组织变白,取组织用多聚甲醛固定进行HE和Masson染色病理学检测纤维化程度差异。
1.2.6 HE染色及Masson染色 HE染色:将小鼠肺组织固定在4%多聚甲醛24 h。在石蜡包埋后,用冰冻切片机将肺组织切成4 μm,先用蒸馏水清洗1~2 s,苏木精液染色(60 ℃)30~60 s,流水洗去苏木精液,1%盐酸乙醇 1~3 s,稍水洗,促蓝液返蓝5~10 s,流水冲洗15~30 s,0.5%曙红液染色30~60 s,蒸馏水稍洗1~2 s,80%、95%和100%乙醇梯度酒精脱水,每步1~2 s,二甲苯浸泡3次,每次 2~3 s,最后用中性树脂封片。
Masson染色:将小鼠肺组织固定在4%多聚甲醛24 h。在石蜡包埋后,用冰冻切片机将肺组织切成4 μm,先用蒸馏水清洗肺切片,再苏木精液染核5~10 min,用蒸馏水洗,再用Masson 丽春红酸性复红液5~10 min,以2%冰醋酸水溶液浸洗片刻,1%磷钼酸水溶液分化3~5 min,直接用苯胺蓝或光绿液染5 min,以0.2%冰醋酸水溶液浸洗片刻,再用80%、95%和100%酒精进行梯度脱色处理,二甲苯浸洗3次,10 min/次,最后使用中性树胶封片。用Masson染色法评价胶原沉积。
1.2.7 羟脯氨酸(Hydroxyproline,HYP)含量检测 肺组织在1.5 ml 5%三氯乙酸中均质化。用蒸馏水洗去质粒,用2 ml的6N HCl在100 ℃水解过夜。用NaOH中和水解产物后,每个样品加入1 ml 0.05 mol/L的氯胺-T溶液,然后在60 ℃下用1 ml 20%对二甲氨基苯甲醛显色20 min。在550 nm处读取吸光度。用纯化的羟脯氨酸制备标准曲线。数据表示为μg羟脯氨酸/mg肺积水。
1.2.8 RIP 以状态良好的HEK-293T细胞为实验对象,稳定过表达NEAT1,将甲醛加入细胞培养液,固定后迅速加入2.5 mol/L甘氨酸解交联,PBS洗涤,刮下细胞,置于离心管中。于冰上将离心管超声,超声后细胞悬液变得澄清透明,离心后取上清液。上清液经稀释后,分成两部分,分别加入特异性抗体和对照IgG,于4 ℃旋转过夜,第2天加入磁珠旋转。分别用稀释缓冲液、低盐溶液、高盐溶液、氯化锂溶液以及TE缓冲液洗涤共7次。用洗脱缓冲液洗脱。将洗脱的产物上样,qPCR检测产物中miR-29a的表达情况。
1.2.9 双荧光素酶报告基因分析 利用Invitrogen法合成了含有野生型(WT)或突变的miR-29a结合位点的lncRNA NEAT1序列,并进行PCR扩增。PCR片段被亚克隆到pMIR-Report质粒(Promega)中荧光素酶基因下游的SacI和HindIII位点。将肺泡上皮细胞接种到24孔板中,培养24 h后,将携带野生型或NEAT1突变的pmirGLO报告载体与miR-29a (100 nmol/L)或miR-NC共转染至肺泡上皮细胞。收集转染24 h后的细胞裂解,并使用双荧光素酶报告试剂盒测定荧光素酶活性。
1.3 统计学分析 采用SPSS 18.0进行统计分析。所有数值都记录为至少3个独立实验的平均SEM。采用双尾t检验来评价各组间差异。P<0.05为差异有统计学意义。
2 结果
2.1 急性肺损伤患者的血清样本中,NEAT1表达上调,miR-29a表达下调 qRT-PCR结果表明,与健康者血清相比,NEAT1在缺血再灌注引起急性肺损伤患者的血清样本表达上调(P<0.001,图1A),而miR-29a在缺血再灌注引起急性肺损伤患者的血清样本表达下调(P<0.001,图1B)。
2.2 低氧低糖诱导的肺纤维化细胞模型中,敲除NEAT1抑制肺泡上皮细胞肺纤维化进程 通过显微镜观察4组细胞的形态,发现模型组相对于对照组细胞形态由鹅卵石型向长梭型变化,而敲除NEAT1可以抑制这种变化(图2A);采用Hoechst染色评估细胞凋亡,与对照组相比,模型组细胞凋亡增加,与模型+sh-NC组相比,模型+sh-NEAT1组细胞凋亡受到抑制(P<0.001,图2B、图2C);经Western blot分析,发现模型组相对于对照组,纤维化标志蛋白col1、col3a1、α-SMA、TGF-β的表达升高,敲除NEAT1可以降低纤维化标志蛋白升高(图2D、图2E);qRT-PCR显示,相对于对照组,模型组NEAT1表达上调和miR-29a表达下调,而敲除NEAT1使miR-29a表达上调(P<0.01,图2F、图2G)。因此,低氧低糖诱导肺泡上皮细胞可以促进细胞凋亡和纤维化,并且使NEAT1表达上调和miR-29a表达下调;敲除NEAT1可以抑制低氧低糖诱导的肺纤维化进程并上调miR-29a表达。

图1 lncRNA NEAT1和miR-29a在缺血再灌注引起急性肺损伤患者血清样本中的表达
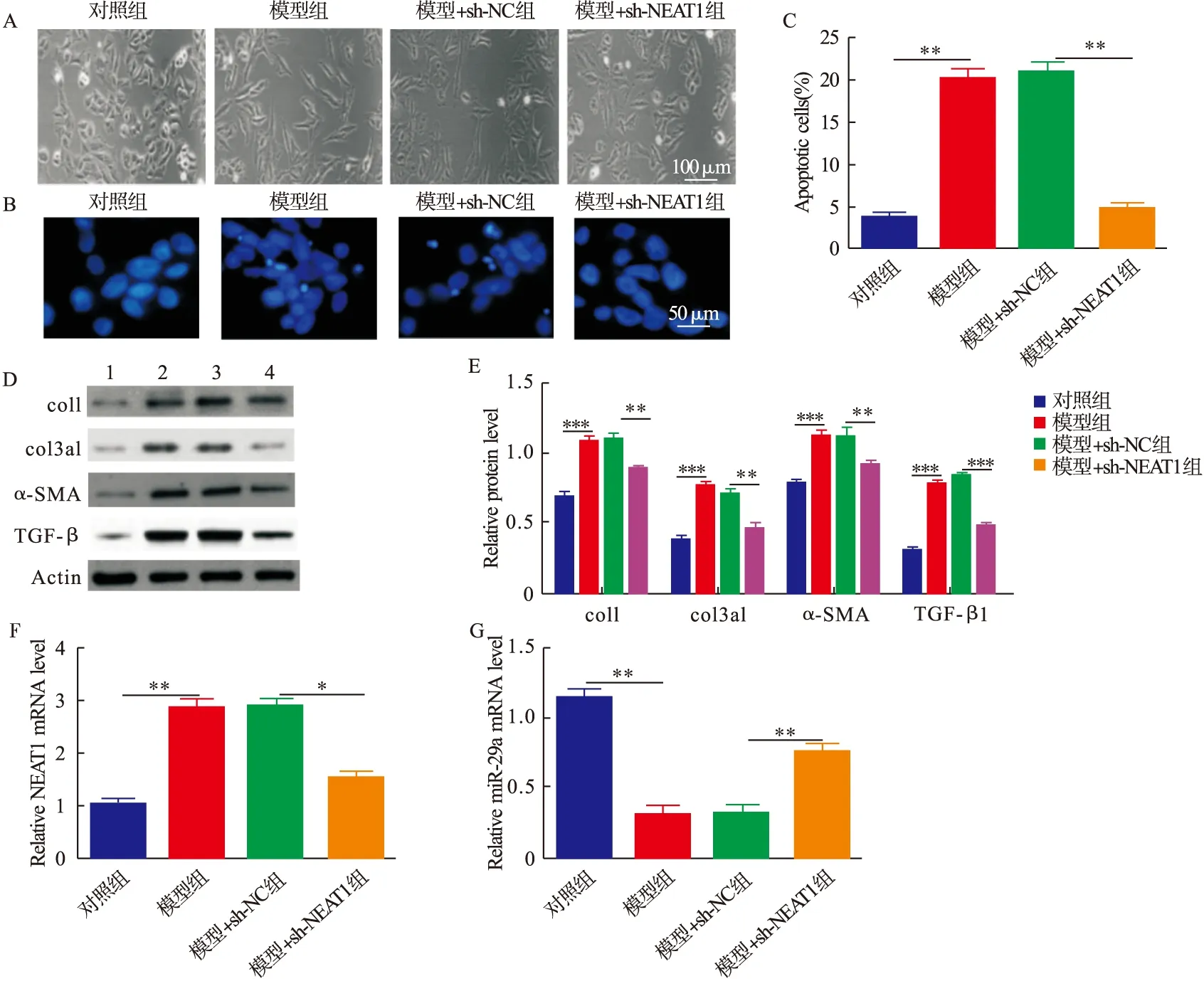
图2 细胞水平敲除lncRNA NEAT1对低氧低糖诱导的肺纤维化进程的影响
2.3 缺血缺氧诱导的小鼠肺损伤动物模型中,敲除NEAT1抑制肺组织纤维化进程 在小鼠体内敲除NEAT1,检测小鼠肺组织纤维化进程以及miR-29a表达。将动物随机分为对照组、模型组、模型+sh-NC组和模型+sh-NEAT1组,小鼠灌流后取肺切片进行H&E和Masson染色,观察组织学变化,发现模型+sh-NEAT1组因缺血缺氧造成的肺纤维化和胶原沉积显著减少(图3A)。肺组织HYP含量检测显示,模型+sh-NEAT1组因缺血缺氧造成的肺纤维化程度显著减轻(P<0.001,图3B、图3C)。Western blot显示,模型+sh-NEAT1组相对于对照组和模型+sh-NC组,肺组织纤维化标志物col1、col3a1、α-SMA和TGF-β表达降低(图3D)。qRT-PCR显示,相对于对照组,模型组NEAT1表达上调,miR-29a表达下调,而敲除NEAT1使miR-29a表达上调(P<0.001,见图3E、图3F)。因此,与体外细胞实验结果一致,体内敲除NEAT1也会抑制肺纤维化相关指标,进而抑制肺纤维化进程,并且上调miR-29a表达。
2.4 NEAT1靶向miR-29a 为了探索肺纤维化中NEAT1与miR-29a的潜在关系,采用TargetScan数据库发现NEAT1与miR-29存在潜在的结合位点(图4A)。采用RIP技术揭示了NEAT1与miR-29a之间的直接结合关系(P<0.001,见图4B)。双荧光素酶报告基因测定揭示NEAT1与miR-29a的靶向关系,发现miR-29a明显抑制了携带lncRNA NEAT1 3′UTR wt的荧光素酶活性,但当报告质粒携带NEAT1 3′UTR mut时,未观察到明显的抑制作用(P<0.01,图4C)。结果显示,NEAT1可以直接与肺泡上皮细胞中的miR-29a结合,且NEAT1可以负调控miR-29a的表达。

图3 体内敲除lncRNA NEAT1对缺血缺氧诱导的肺纤维化进程的影响
2.5 抑制miR-29a逆转了敲除NEAT1对肺纤维化进程的抑制作用 将细胞分为模型+sh-NC组、模型+sh-NEAT1组、模型+sh-NEAT1+anti-miR-NC组和模型+sh-NEAT1+anti-miR-29a组。与模型+sh-NC组相比,模型+sh-NEAT1组细胞形态由长梭型向鹅卵石样变化,细胞凋亡减少,纤维化标志蛋白col1、col3a1、α-SMA、TGF-β的表达降低,miR-29a表达增加;而与模型+sh-NEAT1+anti-miR-NC组相比,模型+sh-NEAT1+anti-miR-29a组细胞形态由鹅卵石样向长梭型变化(图5A),细胞凋亡增加(P<0.001,图5B、5C),纤维化标志蛋白col1、col3a1、α-SMA、TGF-β的表达升高(图5D、图5E),miR-29a表达下降(P<0.001,图5F)。因此,敲除NEAT1抑制肺纤维化进程,而抑制miR-29a逆转了敲除NEAT1对肺纤维化进程的抑制作用。
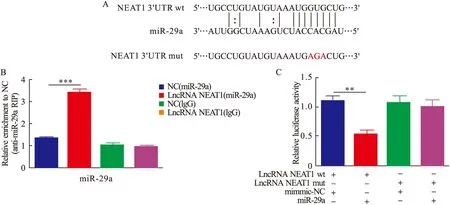
图4 lncRNA NEAT1靶向调控miR-29a

图5 抑制miR-29a对敲除NEAT1后低氧低糖诱导的肺纤维化进程的影响
3 讨论
lncRNA具有调控基因上游启动子区、干扰下游基因的表达,介导染色质重构和组蛋白修饰,与转录因子结合,与特定MicroRNA结合等多种复杂的调控机制[18]。有研究报道,lncRNA在肺纤维化中也有作用,LncH19通过调节miRNA-196a/COL1A1轴介导肺纤维化[19]。通过阻断miR-26a/Smad2来抑制lncRNA PFRL,进而防止肺纤维化[20]。因此,这些lncRNAs可以成为治疗肺纤维化的新靶点。核旁体组装转录因子1(NEAT1)是目前基因组数据集中调控水平最高的lncRNA之一[21]。有研究证实,lncRNA NEAT1在肝纤维化和肾纤维化过程中发挥重要作用[8-9]。但其在低氧诱导的肺纤维化进程中的生物学作用和调控机制尚不明确。本研究发现,lncRNA NEAT1在低氧诱导的肺纤维化组织和细胞中高表达,慢病毒sh-NEAT1转染细胞和感染小鼠时,可以抑制肺泡上皮细胞的凋亡和降低肺纤维化指标蛋白水平,进而抑制低氧诱导的肺纤维化进程。
microRNAs是一类小的非编码核糖核酸,通过抑制RNA翻译来调节基因表达,调节细胞功能[16]。许多MicroRNAs,如miR-21[22]、miR-29[23]、miRNA-145[24]、miRNA-199-5[25]和miR-34a[26]等都在肺纤维化中起关键作用。其中miR-29家族在人肝硬化组织及CCl4和胆管结扎诱导的小鼠肝损伤模型中呈现低表达,且下调miR-29可减少HSCs的活化[27]。Qin等[28]发现,miR-29a可能通过下调DNMT3A抑制心肌成纤维细胞的活化和增殖,从而改善心脏纤维化。但miR-29a对肺纤维化的影响未见有报道。本研究结果显示,miR-29a在低氧诱导的肺纤维化组织和细胞中低表达,通过TargetScan软件预测发现,lncRNA NEAT1与miR-29a可能存在潜在的结合位点,并通过RIP和双荧光素酶报告检测发现lncRNA NEAT1靶向结合,且抑制miR-29a可以逆转敲除lncRNA NEAT1对低氧诱导肺泡上皮细胞的凋亡和降低肺纤维化指标蛋白水平的抑制作用,进而促进了肺纤维化进程。因此,lncRNA NEAT1通过靶向抑制miR-29a促进低氧诱导的肺纤维化进程,然而,此过程中miR-29a下游的靶向基因还不明确。miR-29a常见的靶点包括DNA甲基转移酶(DNMT)1、DNMT3b、COL1A1、AKT1和AKT2等,其中COL1A1 mRNA水平在特发性纤维化疾病中升高[29],提示COL1A1可能作为miR-29a下游的靶向基因来参与lncRNA NEAT1调控的肺纤维化进程。
综上所述,lncRNA NEAT1促进了低氧时肺泡上皮细胞的凋亡和上调了肺纤维化指标蛋白表达,并通过靶向抑制miR-29a促进低氧诱导的肺纤维化进程,为低氧诱导的肺纤维化治疗提供了新的靶点。
猜你喜欢
山东第一医科大学(山东省医学科学院)学报(2022年8期)2022-12-07
中老年保健(2022年2期)2022-11-25
昆明医科大学学报(2022年4期)2022-05-23
昆明医科大学学报(2022年1期)2022-02-28
天津医科大学学报(2021年4期)2021-08-21
现代临床医学(2021年4期)2021-07-31
昆明医科大学学报(2021年4期)2021-07-23
天津医科大学学报(2021年3期)2021-07-21
中华老年口腔医学杂志(2019年2期)2019-04-28
中国民族民间医药·下半月(2011年10期)2011-12-27
