
吉林蛟河不同类型阔叶红松林标记二阶特征
2021-04-10 05:22陈明辉赵中华惠刚盈宋子龙吴相菊
林业科学研究 2021年2期
陈明辉,赵中华,惠刚盈,宋子龙,吴相菊
(1. 中国林业科学研究院林业研究所,北京 100091;2. 国家林业和草原局林木培育重点实验室,北京 100091;3. 北京市植物园,北京 100093;4. 吉林省林业实验区国有林保护中心,吉林 蛟河 132517)
森林结构体现了林木个体 (结构要素)及其属性 (种类、大小、位置) 的连接方式[1],森林结构的异质性可使森林群落中树种多样性增加,促使森林具有更高的稳定性及完整性[2-3],维持森林结构多样性同时被认为是保护生物多样性的最佳途径[4-7]。森林生态系统具有典型的三维特征,组成林分整体的林木个体及其在空间中的排列方式决定了森林系统的属性特征以及功能[8-9],分析群落中种群个体在空间中的分布状态,以及林木个体与其周围相邻木的组织方式、种内种间相互关系以及所处生境的异质性是近年来森林生物物理结构研究的热点之一。
由于环境的异质性、林木间的竞争、种子传播机制等多种因素导致林木位置分布的多样性,采用点格局分析方法可描述林木位置随尺度的变化特征[10-14],如运用Ripley K 函数,O-ring 统计,双相关函数等分析林木的空间分布格局和不同树种的空间相关性,但这些方法大多数仅对林木的位置信息进行了分析,没有包含林木的其它属性,很难与具体的生态学过程相联系。将树种、胸径、树高、冠幅和存活状态等属性标记在林木的位置上,采用标记点格局分析方法即可描述林木属性随尺度变化的特征[15-18],如运用标记相关函数和标记变异函数分析树种、胸径、树高或径向生长在相应尺度上林木属性的相似性和空间自相关性[19-20],可以提供更多的森林空间结构信息。此外,还可以通过构建标记来分析林木间的关系。构建标记通常有差异标记、比例标记、产量标记等,郝珉辉等[20]以林木生长量为标记,运用标记相关函数研究了阔叶红松林林木生长的空间相关性,认为树种生长特征的空间关联格局具有明显的生境依赖性。Pommerening 等[17,21-22]基于标记相关函数的构造原理,提出了标记混交度和标记大小分化二阶特征函数,有效避免了NNSS方法中相邻木选择数量的问题,可解释林分中树种的排列状态和大小分化随尺度变化的情况,同时,对一些生态学过程和假说具有一定的分析和解释能力[23-24],Wang 等[25]运用标记混交度二阶结构特征发现阔叶红松林中大树具有较大的混交度,在一定程度上解释了负密度效应假说。本研究运用单变量双相关函数分析4 块面积为1 hm2的阔叶红松林每木定位样地的林木分布格局特征,然后运用标记双相关函数、标记变异函数、标记混交度二阶特征和标记大小分化阶特征函数联合分析树种和林木大小分化特征,从不同的角度分析不同类型阔叶红松林的空间结构特征,探讨阔叶红松林结构形成的生态过程,以期为进一步保护和培育阔叶红松林,维持生物多样性提供参考。
1 研究区及样地概况
1.1 研究区概况
研究区位于吉林省吉林市蛟河林业实验区管理局东大坡经营区内,43°51′~44°05′ N, 127°35′~127°51′ E,气候属温带大陆性季风山地气候,分布最广的地带性土壤是肥力较高的暗棕壤,植被类型属于温带针阔混交林区域——温带针阔混交林地带—长白山地红松、杉松针阔混交林区,主要针叶树种有:红松 (Pinus koraiensisSieb. et Zucc.)、鱼鳞云杉(Picea jezoensisvarmicrosperma(Lindl))、杉松(Abies holophyllaMaxim) 、 和 臭 冷 杉(Abies nephrolepis(Trautv) Maxim.);主要阔叶树种有:核桃 楸(Juglans mandshuricaMaxim) 、 水 曲 柳(Fraxinus mandshuricaRupr.)、色木槭 (Acer monoMaxim.)、千金榆 (Carpinus cordataBl.)、白扭槭(Acer mandshuricaMaxim.)、紫椴 (Tilia amurensisRupr.)、暴马丁香(Syringa reticulatevar.mandshurica(Maxim. Hara))、蒙古栎 (Quercus mongolicaFisch.)、青楷槭 (A. tegmentosumMaxim)等;常见下木有刺五加(Acanthopanax senticossus(Rupr. et Maxim)Harms)、楔叶绣线菊窄叶变种(Spiraea canescensvar.sublanceolataRehd.) 和胡枝子(Lespedeza bicolorTurcz)、等;主要草本植物有蕨类(Adiantumspp.)、小叶芹(Aegopodium alpestreLedeb.)、苔草(Carexspp.) 、 山 茄 子(Brachybotrys paridiformisMaxim.)、蚊子草(Filipendulaspp.)等。
1.2 样地设置与调查
2007 年5 月到2008 年10 月期间,根据蛟河林业实验区管理局东大坡经营区林班划分情况,在现地踏查的基础上,在试验区建立了4 块面积为100 m × 100 m 的固定样地,运用全站仪对样地内胸径大于5 cm 的林木进行定位,调查林木的胸径、树种、树高等,同时记载林分的郁闭度、坡度、坡向等因子;在样地内四角及中心设置了5 个10 m × 10 m 的小样方,调查样方内的幼苗、幼树情况。在内业分析时,将样地按树种断面积的组成情况依次编号为A、B、C、D(表1),4 块样地代表了4 种类型的阔叶红松林,即样地A 为核桃楸和沙松为主的针阔混交林,样地B 为以核桃楸、水曲柳和红松为主的针阔混交林,样地C 为水曲柳和红松为主的针阔混交林,样地D 为以核桃楸、色木槭和沙松为主的针阔混交林。
2 研究方法
2.1 林木空间分布格局及大小相关二阶特征函数
单变量双相关函数(UPCF)g(r)、标记相关函数(MCF)(r)和 标 记 变 异 函 数(MVF)(r)等3 个函数在以往的研究中已经应用很多[19-20],其中,g(r) 用来分析林木的空间点格局;(r)和(r)用来分析群落标记特征(如林木大小等)的空间相关性特征,二者采用不同的测试函数和归一化方法,(r)以标记属性特征的乘积作为测试函数,即f2(mi,mj)=mimj, 相应的期望值为林分算术平均胸径的平方(μ2);当(r)<1,表明在距离为r的范围内的林木大小属性特征趋向于小于平均值;当(r)>1时 ,距离为r的范围内的林木标记特征趋向于大于平均值;而则表达标记特征随着尺度的变异程度,其测试函数为当时 ,表明在距离为r的范围内相似大小林木有聚集的特征;当时,表明在距离为r的范围内相似大小林木间排斥,而不同大小的林木聚集。

表1 4 块样地林分基本特征Table 1 The basic characteristics of fourdifferent samples
2.2 标记大小分化度及标记混交度二阶特征
Pommerening 等[21]基于标记相关函数的构造原理,以Gadow 提出的简单混交度及大小分化度分别作为测试函数[26],其中,标记大小分化度二阶特征测试函数表达式为:
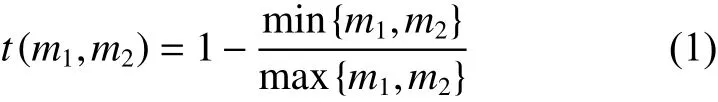
式中,m1和m2是标记在两个不同林木位置上的林木大小定量属性值,由于林木胸径测量容易且能够稳定代表林木的大小属性特征,因此本研究中以林木胸径值计算。林分中每株树可作为参照树,将其胸径值(m1) 与其距离为r的相邻木(m2)进行比较,可构建标记大小分化二阶特征检验函数:
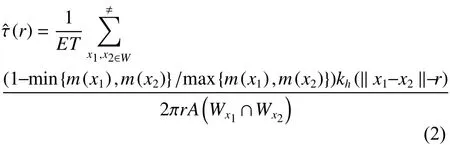
式中,x1和x2是在所观测的窗口中两个任意的点。k为Epanechnikovkenel 核 函 数,A(Wx1∩Wx2)为Wx1与Wx2相交面积,用于矩形样地的边缘矫正。当标记大小分化度(r)=1时,表示群落中林木大小随机分布;当(r)>1时,表示群落中不同大小林木个体聚集或相似大小林木排斥;当^ τ(r)<1时,表示群落中相似大小林木个体聚集或不同大小林木个体排斥。ET为期望大小分化度,其计算过程首先将样地中林木胸径由小到大进行排序:然后定义一个累加变量Di,其表达式为:
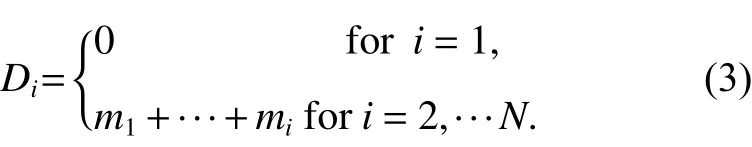
根据样地林木数量N及上式中的Di可得到林分期望大小分化值为:
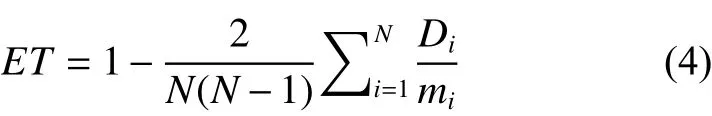
同样,将混交度的指示函数作为测试函数可构建标记混交度二阶特征。其中测试函数表达式为:

式中,m1和m2是标记在两个不同位置上的林木树种属性值。式(5)为指示函数,当m1和m2为不同树种时,Mi值为1,否则其值为0。
标记混交度二阶特征检验函数表达式为:
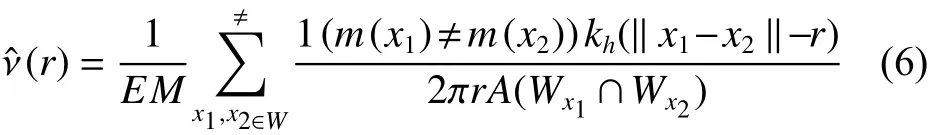
式中,x1、x2是在所观测的窗口中两个任意的点。A为 边缘矫正权重,kh为Epanechnikovkenel核函数。EM为期望混交度,因此当标记混交度的(r)的值等于1 时,表达了群落中的树种间随机分布;当(r)>1时,表达了不同树种聚集或同种排斥;当时,为相同树种聚集或不同树种间负相关。其中,林分的期望混交度计算为:
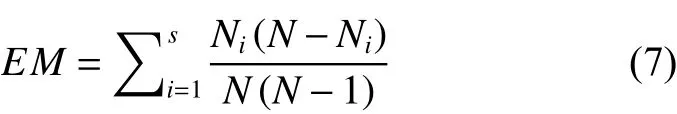
其中s为树种数,N为样地林木株数,Ni为第i个树种的林木个数。
2.3 零模型检验及数据计算
本研究在进行二阶特征乘积密度估计时采用Epanechnikovkenel 函数(kh),其中kh的带宽既不能设置的太窄,也不能太宽,太窄时包含在圆环中的点太少,会造成随机波动,而太宽时包含太多的点,则会忽视了所关注的尺度距离r处的特征。本研究根据样地的密度将带宽统一设置为2 m。在进行林木分布格局分析时,采用完全空间随机过程(CSR)均质泊松过程作为零模型;在对标记胸径二阶特征分析时,采用随机标记零模型,即保持林木的位置坐标不变,将标记属性随机分配到林木的位置上[17]。在进行标记混交度二阶特征分析时,采用了随机叠加或种群独立性检验[17,21,27];对于多树种混交的林分而言,设置一组树种的位置固定不变,移动另一组树种,两组树种林木个数约为总数的一半。对于标记大小分化二阶特征分析时仍随机标记零模型。运用蒙特卡洛完全随机分布模型重复1 000 次,模拟产生95%的置信区间,并绘制上限和下限分别为2.5%和97.5%的包迹线,对标记函数结果偏离随机状态的进行显著性检验。
在数据计算和分析时,均使用R 统计软件进行。标记双相关函数、标记变异函数、标记混交度和标记大小分化度相关代码文件可在Pommerening森林生物实验室(Pommerening's Forest Biometrics Lab)获得。为避免边缘效应对结果的影响,采用NN1 方法进行边缘校正[28]。
3 结果与分析
3.1 不同类型阔叶红松林林木分布格局
单变量双相关函数g(r) 结果表明(图1),4 块样地中的林木分布格局存在一定的差异。样地A 中的林木在r= 5~8 m 的尺度上和r大于10 m的尺度上,林木呈现显著聚集的格局特征;样地B 中林木的分布格局整体上为随机分布;样地C 和样地D 分别在r= 8~11 m 和r= 7~12 m 的尺度上呈现显著聚集分布的趋势,而在其他尺度上林木的分布格局为随机分布。

图1 样地林木分布格局随尺度变化Fig. 1 Spatial distribution pattern of trees with scale
3.2 不同类型阔叶红松林标记胸径二阶特征
3.2.1 不同类型阔叶红松林标记胸径相关函数特征 以胸径为标记的相关函数表明(图2 左),样地A中的林木在r= 0~50 m 的观察尺度上,标记胸径相关函数(r)的观测值都小于1,同时在r=0~8 m 和r= 13~33 m 的尺度上小于随机标记零模型,表明成对相邻木具有小树特征;样地B、C 和D 标记相关函数结果非常类似,即(r)的观测值在小尺度上小于1,随着研究尺度的增加,(r)的观测值在1 附近波动,说明这几个样地中的林木在小尺度上表现出相邻木多为小树,其中,样地B、C 和D 分别在r= 0~8 m,r= 0~7 m 和r= 0~6 m 的尺度上较为显著,而在其它尺度上林木的胸径大小随机分布。
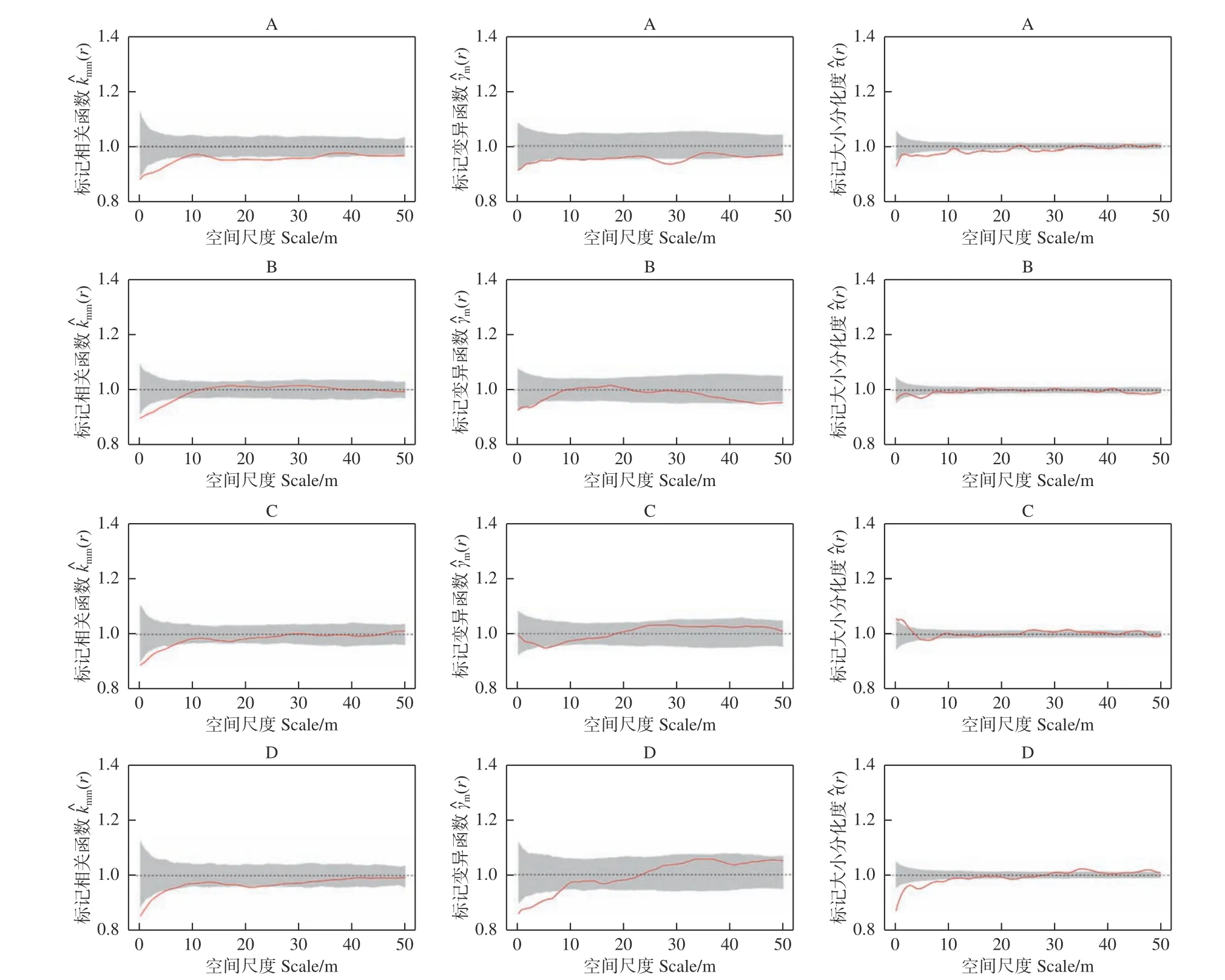
图2 不同类型阔叶红松林胸径标记二阶特征Fig. 2 Second-order characteristics of DBH markers in different types of broad-leaved Korean pine forests
3.2.2 不同类型阔叶红松林胸径标记变异特征 标记胸径变异函数结果表明(图2 中),样地A 中的林木在r= 0~50 m 的观察尺度上,(r)的观测值均都小于1,说明样地A 中的林木在对应尺度上林木具有相似胸径聚集的趋势,随机标记零模型检验表明,在r= 25~33 m 的尺度上这种趋势较为显著。样B 和样地C 中的林木在r= 0~50 m 的观察尺度上,标记胸径变异函数(r)的观测值始终变化在1 的附近波动,完全落入了包迹线内,表明这2 块样地内林木胸径大小相关性不明显;样地D 内的林木在标记胸径变异函数(r)的观测值在小尺度上小于1,随着尺度的增加趋向于在1 的附近波动,样地D 内的林木在r= 0~9 m 的尺度上呈现显著的空间正自相关,即相似胸径大小的林木聚集分布,而在其它尺度上林木胸径大小呈现空间上不相关的特征。
3.2.3 不同类型阔叶红松林标记大小分化二阶特征 不同类型阔叶红松林标记大小分化二阶特征函数表明(图2 右),4 块样地中的林木大小分化特征并未完全落在零模型内,说明4 块样地内不同大小林木的分布并非是完全随机分布的格局。其中,样地A 内林木的标记大小分化二阶特征函数(r)观测值在不同尺度下小于1,说明林木胸径的大小分化度显著小于期望大小分化度,相似大小的林木聚集在一起;样地B 和D 样地内林木的标记大小分化度二阶特征表现出类似的趋势,即这2 块样地仅在小尺度上表现出相似大小林木显著聚集的特征,对应的尺度分别为r= 4~6 m 和r= 0~12 m;同时样地D 内林木的标记大小分化度在r= 0~12 m的尺度上观察值与期望值相差较大。与其他样地不同,样地C 内林木的标记大小分化度二阶特征函数(r)的值在r= 0~5 m 的尺度上大于1,表明林木胸径的大小分化度大于期望大小分化度,意味着不同大小的林木聚集在一起,即小树与大树吸引,小树与小树排斥。
3.3 不同类型森林标记混交度二阶特征
根据随机叠加(或种群独立性)零模型(图3)结果表明,4 块样地内的树种整体分布属于完全随机分布,标记混交度二阶函数(r)的观测值在一定的尺度范围内均小于1,并且随着研究尺度的增加,其值逐渐接近于1,说明各样地林木的树种在相应的尺度上存在同种聚集的现象,随着尺度的增加,样地中林木的树种分布逐渐趋于随机分布。此外,从各样地的标记混交度二阶特征函数值还可以看出,各样地(r)值在小于1 的范围内曲线变化总体趋势不同,样地A 和C 的(r)值在相应的尺度上从最小值逐渐增加到接近于1,样地B 和样地D 的(r)值在相应的尺度上先迅速增加,然后平缓地接近1,说明不同类型的阔叶红松林尽管都在一定尺度上存在着相同树种聚集的现象,但其范围和变化趋势存在明显差异。
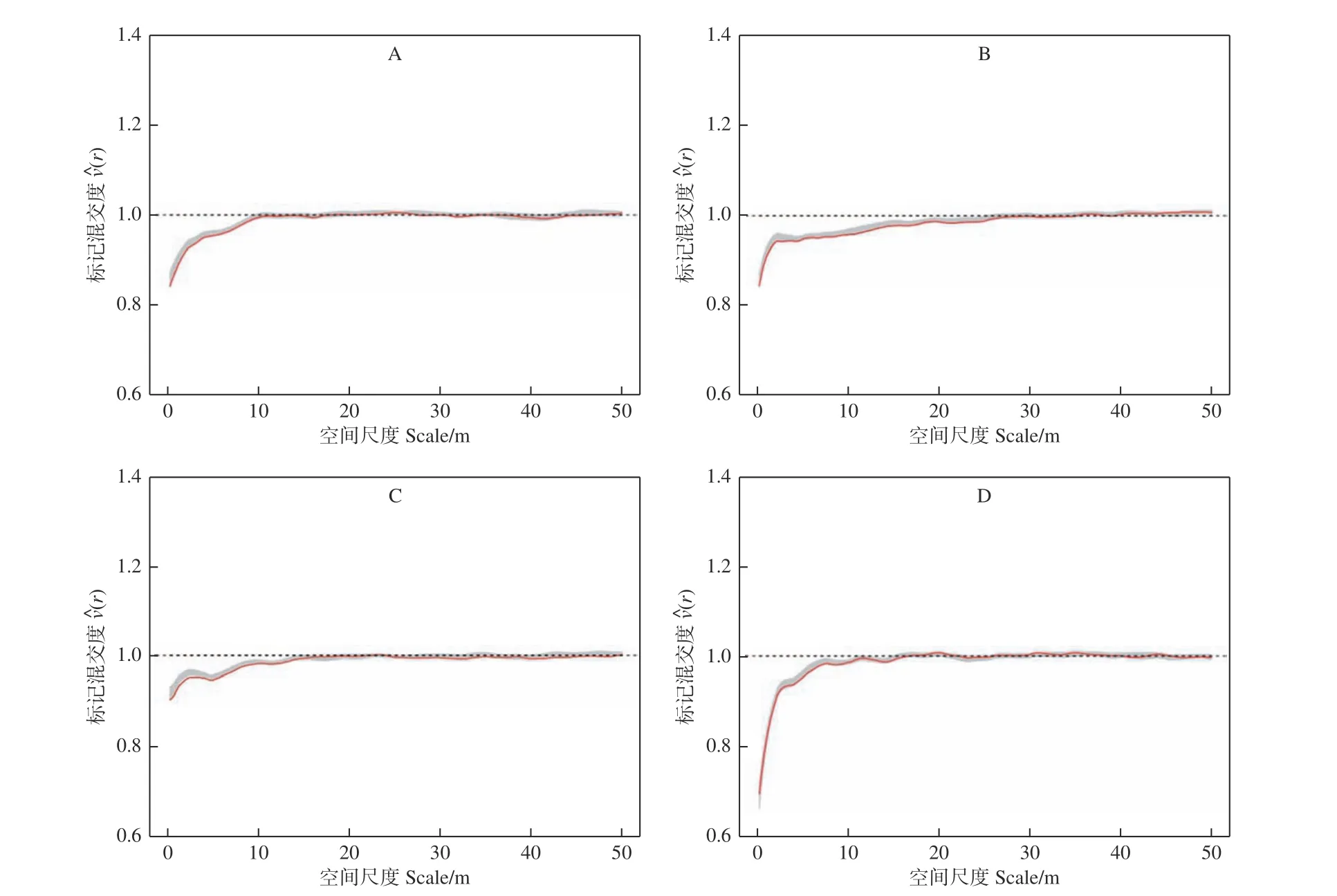
图3 不同类型标记混交度二阶特征Fig. 3 The second-order characteristics of mark mingling in different types of Korean pine broad-leaved forest
4 讨论
本研究分析了不同类型阔叶红松林林木分布格局和标记二阶结构特征,发现不同类型的阔叶红松林的林木分布格局、大小分化标记二阶特征差异明显,特别是水曲柳红松林在小尺度上表现出了明显的小树与大树吸引,小树与小树排斥的特征,明显不同于其他3 个类型,这可能与红松阔叶林的发育阶段及干扰程度有关。在4 种阔叶红松林中,水曲柳红松林中的红松比例最高,达到了2 成,根据李景文的研究,红松阔叶林中阔叶树的伴生作用主要发生在红松发育的前期和中期,在这一范围内,红松的株数极少;随着红松的发育,阔叶树逐步退出群落或者是长寿命的阔叶树与红松之间经过竞争,逐渐达到一种稳定的状态,从而在小尺度上形成大树与小树聚集分布的特征[29]。阔叶红松林的标记混交度特征表明不同类型的阔叶红松林树种分布整体上呈现随机分布,但在小尺度上存在同种聚集的现象,其范围和变化趋势存在明显差异,这进一步说明了不同类型的阔叶红松林处于不同的发育阶段,相同树种聚集程度较高的林分还处于发育的早期阶段,更新主要发生在母树周围;而聚集程度较低的林分是演替的结果,相同树种已经发生自疏现象,其他树种的种子由不同媒介传播,并在一定的范围内更新。根据分析结果可以推测,水曲柳红松林可能处于红松发育的中期或者在历史上受到的干扰程度较小,而其它3 种类型的阔叶红松林处于发育的早期或者在历史上受到的干扰程度较大,林木间的自疏作用尚未完成。此外,种内和种间关系、环境的异质性,如水分、土壤、温度、小地形等外部因素也可能是造成林木大小或树种分布呈现不同分布格局的原因,还需要进一步对不同类型阔叶红松林标记二阶特征的形成进行深入探讨和研究。
阔叶红松林是我国东北林区的特有森林类型,在温带针阔混交林类型中占重要地位[30-32]。现有红松阔叶林大多为遭受不同程度的人为干扰后恢复形成的次生林,如何加快红松阔叶林恢复是经营中面临的一个主要问题。邓守彦等认为红松阔叶林在干扰过后,完全可以通过70 年的天然恢复接近原始林群落水平,然而,天然林自然恢复是一个漫长的过程,如果辅以合理的人工措施,必然会加速红松阔叶林的恢复[33]。本研究采用的标记二阶特征函数可以对林分不同尺度上的树种和林木大小的分化特征进行细化,其结果对红松阔叶林加速恢复有指导作用,即对于同种聚集程度较高且相同大小林木聚集的林分,可以选择红松优良个体作为培育对象,通过调整其周围较其大的伴生阔叶树种,创造林窗,为红松生长提供充足的营养空间;而对于相同树种聚集且不同大小林木聚集的林分,则主要是调整同种间的资源竞争,伐除相同伴生树种的小树,促进林下红松的更新。当然,在对处于不同恢复阶段的红松阔叶林经营时,还要考虑林分整体的稳定性,需要针对具体林分进行全面的分析。
5 结论
不同类型阔叶红松林的林木分布格局和林木大小分化特征差异明显,树种分布整体上呈现随机分布,但在小尺度上存同种聚集的现象,其范围和变化趋势存在明显差异。不同测试函数标记二阶特征联合分析进一步细化了林木大小分化特征,能够表达处于不同发育阶段阔叶红松林的结构特征,对阔叶红松林加速恢复经营具有一定的指导意义。
猜你喜欢
湖南林业科技(2022年4期)2022-08-29
中等数学(2021年9期)2021-11-22
陕西农业科学(2021年10期)2021-11-09
防护林科技(2021年3期)2021-09-12
中国房地产业·上旬(2021年4期)2021-04-27
国际木业(2018年4期)2018-09-10
数学学习与研究(2017年20期)2018-01-02
前卫文学(2016年3期)2016-07-01
中学数学杂志(初中版)(2016年3期)2016-06-24
湖南师范大学学报·自然科学版(2014年3期)2014-10-24
