
mgrB和mcr-1在耐多黏菌素猪大肠杆菌中的作用
2021-04-12 09:15邝启红李文娅李垠树贾雅婷胡功政河南农业大学动物医学学院河南郑州450046
中国兽医学报 2021年3期
邝启红,李文娅,李垠树,贾雅婷,胡功政,苑 丽 (河南农业大学 动物医学学院,河南 郑州 450046)
多黏菌素是一种广谱抗革兰阴性菌的阳离子多肽,主要用于治疗多重耐药革兰阴性菌感染,尤其是产碳青霉烯酶肠杆菌科细菌导致的感染。近年来,耐多黏菌素的肠杆菌科细菌日益增多,导致人医和兽医临床防控耐药革兰阴性菌感染更加困难。目前已阐明多黏菌素的耐药机制主要有2种:染色体介导的双组分信号转导系统PmrAB和PhoPQ的表达量升高及其上游阻遏蛋白MgrB的特异性突变和质粒介导的多黏菌素耐药基因mcr[1-2]。mcr基因具有磷酸乙醇胺转移酶活性,细菌产mcr后会导致菌体内磷酸乙醇胺合成增多,从而修饰类脂A,并最终导致细菌对黏菌素耐药[1]。MgrB是一种47个氨基酸编码的跨膜蛋白[3],是MgrB-PhoPQ-PmrHFIJKLM信号通路中的负性调控蛋白[4-5],可通过抑制PhoQ组氨酸激酶的活性,阻遏磷酸基团转移给PhoP,从而抑制下游操纵子的表达[6]。近来有研究表明:mgrB基因的缺失、变异、插入或截断失活以及mgrB启动子区突变或插入失活,都会导致PhoPQ系统和pmrHFIJKLM操纵子过表达[7-9],进而造成细菌对多黏菌素耐药。
细菌的mgrB基因变异或携带mcr基因都会导致其对多黏菌素耐药,但目前尚不清楚在耐多黏菌素大肠杆菌中这两种机制存在的差异。本试验对收集的165株猪源大肠杆菌采用微量肉汤稀释法测定多黏菌素的敏感性,并对耐药菌株的mcr-1~mcr-5基因以及mgrB基因进行检测及测序比对分析,进而评估mgrB和mcr-1在大肠杆菌对多黏菌素耐药的贡献,为防控多黏菌素耐药的传播和延缓耐药趋势提供试验数据和依据。
1 材料与方法
1.1 菌株和质粒165株大肠杆菌由本实验室于2016—2017年自安徽、河南、湖南、湖北、江西、山西、陕西7省不同地区猪病料样本(肠、肺脏、淋巴、脾脏、肝脏等)和健康猪肛门拭子采集分离保存;大肠埃希菌ATCC25922(购自中国普通微生物菌种保存中心)为质控菌;pBAD/HisA质粒(由本实验室保存)具有氨苄西林抗性。
1.2 主要试剂和药品LB肉汤、LB琼脂等培养基均购自北京奥博星生物技术有限责任公司;质粒小提试剂盒和DNA胶回收试剂盒购自美国OMEGA公司;Primer STAR高保真酶,XholⅠ和Hind Ⅲ内切酶,T4DNA连接酶和荧光染料Mix等均购自TaKaRa公司;DL2000、DL5000 Marker为北京擎科新业生物科技有限公司产品;氨苄西林钠购自北京索莱宝科技有限公司;多黏菌素(含量90%)是由河南牧翔动物药业有限公司提供。
1.3 菌株复苏取甘油保存的菌种并充分振荡,按1∶100比例无菌接种于3 mL新鲜的LB肉汤,置于37℃振荡培养12 h,用无菌接种环蘸取一环菌液划线接种于LB琼脂平板上,倒置于37℃温箱中培养12 h,后挑取单个菌落接种于LB肉汤,37℃振荡培养12 h后备用。
1.4 质粒提取pBAD/HisA质粒的提取按照美国OMEGA公司的质粒小量提取试剂盒说明书进行,将收集的液体置于-20℃保存。
1.5 药物敏感性测定
1.5.1药液的配制 用分析天平称取多黏菌素和氨苄西林钠适量,无菌配制为5 120 mg/L,0.22 μm滤膜过滤分装至1.5 mL EP管中,置于-20℃保存备用。
1.5.2MIC值测定 采用微量肉汤稀释法[10]测定多黏菌素对165株猪大肠杆菌的最小抑菌浓度(minimal inhibitory concertration,MIC),结果按照美国临床实验室标准化协会(CLSI)标准判读,大肠杆菌ATCC25922为质控菌。
1.6 多黏菌素耐药基因的检测
1.6.1菌株DNA提取 采用常规煮沸法[11]提取各菌株的DNA,-20℃保存备用。
1.6.2引物设计 根据GenBank公布的mgrB核苷酸序列,应用primer 5.0分析软件设计引物,mcr-1~mcr-5引物序列参考相关文献,由上海生物工程有限责任公司合成。其中,mgrB-F/mgrB-R包含mgrB基因及其上游启动子区,引物序列见表1。

表1 多黏菌素耐药基因PCR扩增的引物序列
1.6.3多黏菌素耐药基因比对分析 PCR检测并测序分析耐药猪源大肠杆菌的mcr-1~mcr-5流行情况。mgrB基因全序列及其启动子区经PCR扩增后,产物经胶回收、纯化后,用T4DNA Ligase 4℃过夜连接至pBAD/HisA质粒,连接产物用化学转化法转入DH5α感受态细胞,用引物mgrB-F/mgrB-R进行PCR验证,并提取重组质粒经XhoⅠ和Hind Ⅲ双酶切鉴定,送至上海生物工程股份有限公司测序,测序结果用DNAStar软件分析,以大肠杆菌K-12 MG1655为参照菌,进行同源性比对分析。
2 结果
2.1 猪大肠杆菌对多黏菌素的敏感性测定多黏菌素对165株猪大肠杆菌的MIC值分布见图1,其中147株(89.1%)对多黏菌素耐药,MIC的范围为4~32 mg/L,且MIC=16 mg/L时菌株数最多,有82株。MIC值≤1 mg/L共有18株,敏感率为10.9%。
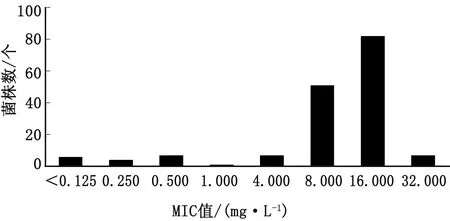
图1 165株猪源大肠杆菌对多黏菌素的MIC值分布
2.2mcr耐药基因的鉴定PCR检测耐多黏菌素猪大肠杆菌的mcr-1~mcr-5基因。结果显示,147株耐多黏菌素的猪大肠杆菌中除EPF25菌未检出mcr基因外,其余146株菌都仅携带mcr-1基因(图2),所有菌株均未检出mcr-2~mcr-5基因。
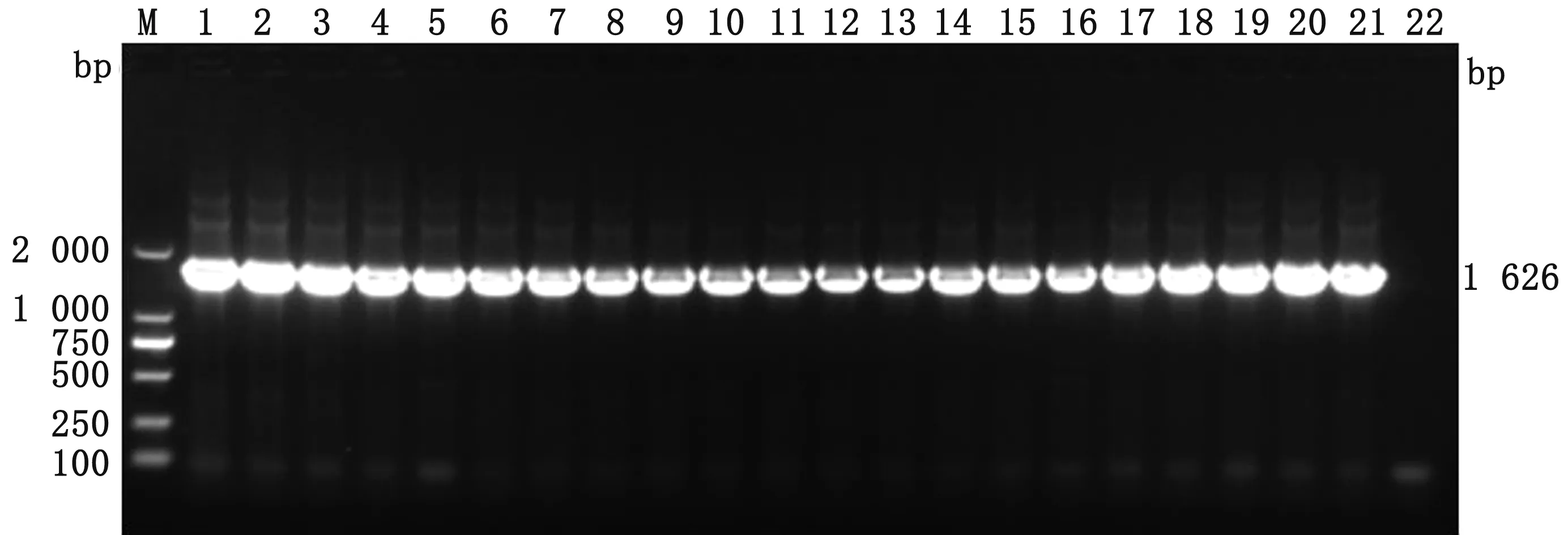
M.DL2000 DNA Marker;1~20.部分多黏菌素耐药菌;21.阳性对照;22.阴性对照
2.3mgrB基因的突变将mgrB基因及其启动子区连接pBAD/HisA载体并转入感受态细胞DH5α后,提取重组质粒,PCR鉴定扩增出预期大小的片段并经XhoⅠ和HindⅢ双酶切证实为mgrB-pBAD/HisA重组质粒(图3)。
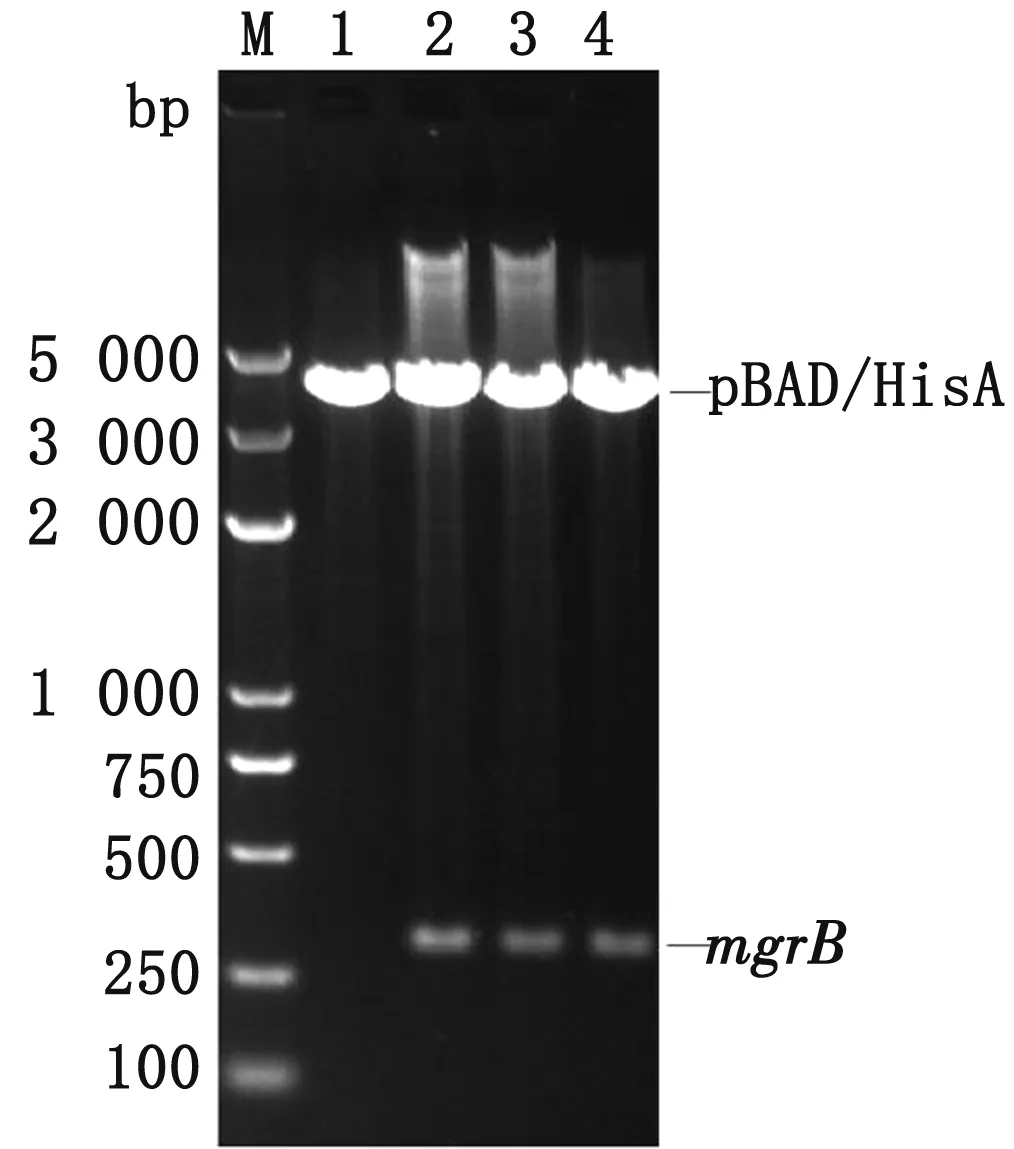
M.DL5000 DNA Marker;1.pBAD/HisA;2~4.pBAD/HisA-mgrB
以大肠杆菌K-12MG1655(GenBank号:NZ_CP014348.1)为参照菌株,mgrB基因经PCR检测并测序比对分析。结果表明,mgrB启动子区变异的有104株(70.7%,104/147),且均携带mcr-1基因,其中99株菌有相同的碱基突变位点,均为mgrB编码区上游12 bp处的A→G碱基;3株菌(E176、E186、E224)含2个碱基突变位点:编码区上游12 bp 处的AG碱基和27 bp处的C→T碱基突变;E193菌(编码区上游44 bp处G→A碱基突变)和EPD04菌(编码区上游38 bp处C→G碱基突变)仅含1个突变位点,且与上述不同。mgrB编码区氨基酸突变的有11株(0.07%,11/147),其中仅有EPF25菌不含mcr基因;有6株菌(EPA05、EPA06、EPA09、EPA25、EPA27和EPD11)均为33位的谷氨酰胺突变为精氨酸,有3株菌(E8、E180、E221)是31位的天冬氨酸突变为甘氨酸,EPD23菌为33位的谷氨酰胺突变为组氨酸,EPF25菌为42位的天冬酰胺突变为丝氨酸。代表性菌株mgrB基因及其启动子区序列已提交至NCBI,获得的GenBank序列号见表2。比较多黏菌素对受试菌的MIC值及耐药菌mcr-1基因、mgrB变异之间的关系发现它们之间的相关性不明显,猪大肠杆菌对多黏菌素耐药是由于mcr基因和mgrB变异共同作用的结果。

表2 耐多黏菌素的大肠杆菌mgrB(与K-12MG1655相比)的突变位点
3 讨论
近年来,mgrB基因的突变、缺失或者插入序列在多黏菌素耐药肺炎克雷伯菌中已有报道,且已被证明在肺炎克雷伯菌黏菌素耐药中起重要作用[4,14-15]。大肠杆菌多黏菌素耐药机制的相关研究主要集中于质粒介导的mcr基因[8,16]。肺炎克雷伯菌的mgrB启动子区插入类IS903序列可引起mgrB表达量显著下降,从而导致PhoPQ系统和pmrHFIJKLM操纵子过表达,最终使该菌对多黏菌素表现耐药[17]。同时,OLAITAN等[18]在患者中分离的耐多黏菌素肺炎克雷伯菌中的mgrB启动子区也发现IS903插入序列。HAEILI等[19]在mgrB启动子区发现IS插入序列。但有关mgrB启动子区碱基突变的相关报道尚未见。本研究在多黏菌素耐药的147株菌中检测到104株mgrB启动子区碱基突变,推测mgrB启动子区突变可能会导致mgrB基因表达减少,进而导致PhoPQ系统上调,降低大肠杆菌对多黏菌素的敏感性,但尚须要进一步研究证实。
本试验结果表明有11株耐多黏菌素猪大肠杆菌的mgrB编码区氨基酸发生突变,且以往研究已证明这3处氨基酸突变与细菌对多黏菌素耐药相关。EPF25菌的mgrB基因发生N42S突变,该突变与DAGHER等[20]在黎巴嫩医院中分离的耐多黏菌素阴沟肠杆菌的mgrB基因中的突变相同;2017年,POIREL等[21]也在耐多黏菌素动物源肺炎克雷伯菌中的mgrB中发现了N42Y突变。2019年,HUANG等[22]从中国食用动物收集的大肠杆菌分离株中检出mgrB基因的D31G突变,该突变也同样发生在本试验的E8、E180和E221等3菌株中。2019年,KIM等[23]在韩国猪源大肠杆菌mcr基因阴性分离株中检出mgrB的Q33R突变,而本试验在大肠杆菌EPA05、EPA06、EPA09、EPA25、EPA27和EPD11等6株菌中检出存在同样的氨基酸突变。推断mgrB基因的31、33和42位的氨基酸突变与细菌对多黏菌素产生耐药密切相关。
2018年,ZHANG等[24]发现当肺炎克雷伯菌中因mgrB失活而呈现对多黏菌素高度耐药后,既使该菌株再获得mcr-1基因也不会引起其对多黏菌素耐药程度的改变,故猜测失活的MgrB可能掩盖了mcr-1基因的功能。而齐小梅等[7]在大肠杆菌耐药菌中未检测到mgrB突变,并认为大肠杆菌对黏菌素耐药的主要原因是获得mcr-1基因。本试验比较多黏菌素对受试菌的MIC值及耐药菌mcr-1基因、mgrB变异之间的关系发现它们之间的相关性不明显,说明猪大肠杆菌对多黏菌素耐药是由于mcr基因和mgrB变异共同作用的结果。
猜你喜欢
化学工程师(2022年1期)2022-02-23
教学考试(高考生物)(2020年6期)2020-11-23
广东医科大学学报(2020年4期)2020-08-24
食品与生物技术学报(2020年8期)2020-01-06
科学24小时(2019年5期)2019-06-11
发明与创新(2019年9期)2019-03-26
天然产物研究与开发(2018年9期)2018-10-08
大连海洋大学学报(2018年3期)2018-07-24
China Communications(2017年12期)2017-04-10
广东饲料(2016年6期)2016-12-01
