
转基因白桦杂交子代种子活力及外源基因遗传规律分析
2021-04-19 08:35吕东林郭译文黄海娇
植物研究 2021年4期
吕东林 李 腾 郭译文 姜 静 黄海娇*
(1. 林木遗传育种国家重点实验室,东北林业大学,哈尔滨 150040;2. 辽宁林业职业技术学院,沈阳 110101)
聚合育种是指利用杂交育种、现代分子育种等途径实现多个外源基因聚合,从而达到育种目标的方法。通过多基因聚合等分子育种手段以培育高产、优质、多抗、高效、稳定的农作物新品种,已成为国内外学者研究的热点[1]。迄今为止,已在多种农作物上得到应用,Hittalmani 等[2]将稻瘟病抗性基因Piz5、Pi1、Pita 成功聚合到BLI24 中,巴 沙 拉 特 等[3]成 功 获 得Xa4、Xa5、Xa13、Xa21 4 个抗白叶枯病基因的水稻子代纯合体;董娜等[4]实现了小麦抗白粉病基因Pm21 和Pm13 的聚合育种;Matsumoto 等[5]对黄瓜(Cucu⁃mis sativus)进行过基因聚合能够提高其抗性和抗谱的研究报道,朱明涛等[6]对番茄(Lycopersicones⁃culentum Mill.)聚合育种的研究得出了一致结论。
白桦(Betula platyphylla Suk.),落叶乔木,主要分布于我国北方,具有分布广、耐低温等特点[7~8]。白桦树皮灰白色,可成层剥裂,树干姿态优美,具有独特的观赏价值,常被用作行道树、庭院树等[9]。研究人员采用基因工程育种手段相继对白桦进行了遗传改良,获得了童期缩短的BpAP1 转基因白桦[10],提高抗旱耐盐性的TabZIP 转基因白桦[11],促进木质素含量降低的BpCCR 转基因白桦[12],维持吲哚乙酸(IAA)、茉莉酸(JA)等动态平衡的BpGH3.5 转基因白桦[13~14],抗低温的BpMYB4转基因白桦[15],并对能够影响白桦生长发育和抗性的BpTCP7 和BpTCP8 基因也进行了研究[16~17]。然而,有关转基因白桦聚合育种的研究却鲜见报道,陈素素等[18]以TabZIP、BpAP1 转基因白桦为试材,进行常规育种,研究表明目的基因通过雌配子的传递效率高于雄配子,雌、雄配子外源基因传递效率分别在60.00%、30.00%左右,并获得了TabZIP、BpAP1 基因聚合的PB1、PB5 白桦T1代以及单个BpAP1 基因的P13、P15 和P20 白桦T1代。赵洁等[19]对转入Cry1Ac 和Cry3A 基因的双Bt转基因巨霸杨(Populus deltocdes 50×P. deltocdes 36)进行过研究报道。本文在陈素素等研究的基础上,以BpAP1 转基因白桦T1代、BpAP1TabZIP 转基因白桦T1代、BpGH3.5 和BpCCR 转基因白桦为杂交亲本,设计杂交组合,拟进一步验证转基因白桦目的基因的遗传规律,期望获得3 个基因聚合的T2代个体,是对白桦转基因聚合育种的后续报道。
1 材料方法
1.1 试验材料
1.2 种子活力的测定及子代生长性状分析
1.2.1 种子千粒质量的测定
采用百粒法对白桦种子的千粒质量进行测定,从各家系中分别随机取800 粒种子,设置8 个重复,每个重复100 粒,各重复分别称重(mg),小数点后面保留两位小数,计算各家系白桦种子的千粒质量。
1.2.2 种子活力指标的测定
参照陈素素等[18]测定种子活力的方法进行种子发芽实验,每个家系重复3 次,每个重复100 粒种子,发芽试验结束后分别计算发芽率、发芽势、发芽指数等指标。随后,进行播种育苗,并于2017年4月初和9月末分别调查15个家系的T2代苗高,并运用以下公式计算苗高生长量和苗高相对生长量:
苗高生长量=9月末苗高-4月初苗高
苗高相对生长量=苗高生长量/4月初苗高
1.3 观察指标 观察并记录行CPT1周内,CPT前1 h(T1)、CPT开始初期1 h(T2)、CPT结束后1 h(T3)这3个时间节点患者的呼吸频率(RR)、氧合指数(OI)、血氧分压(PaO2)及血氧饱和度(SaO2),同时观察记录血流动力学指标:心率(HR)及平均动脉压(MBP)。
1.2.3 数据处理
运用Microsoft Excel 2007 进行数据处理并对百分数据进行反正弦转换,利用SPSSv22.0 软件进行方差分析、多重比较和相关性分析。
家系遗传力(H2)公式:
式中:F为方差分析中的F值。
1.3 外源基因的分子检测
1.3.1 白桦总DNA的提取
采用植物基因组DNA 提取试剂盒(天根生化科技(北京)有限公司,Cat.#DP321-03),分别选取G7、G8、G22、C13 等4 个家系,每个家系随机采取12~31株幼苗的叶片进行总DNA的提取。
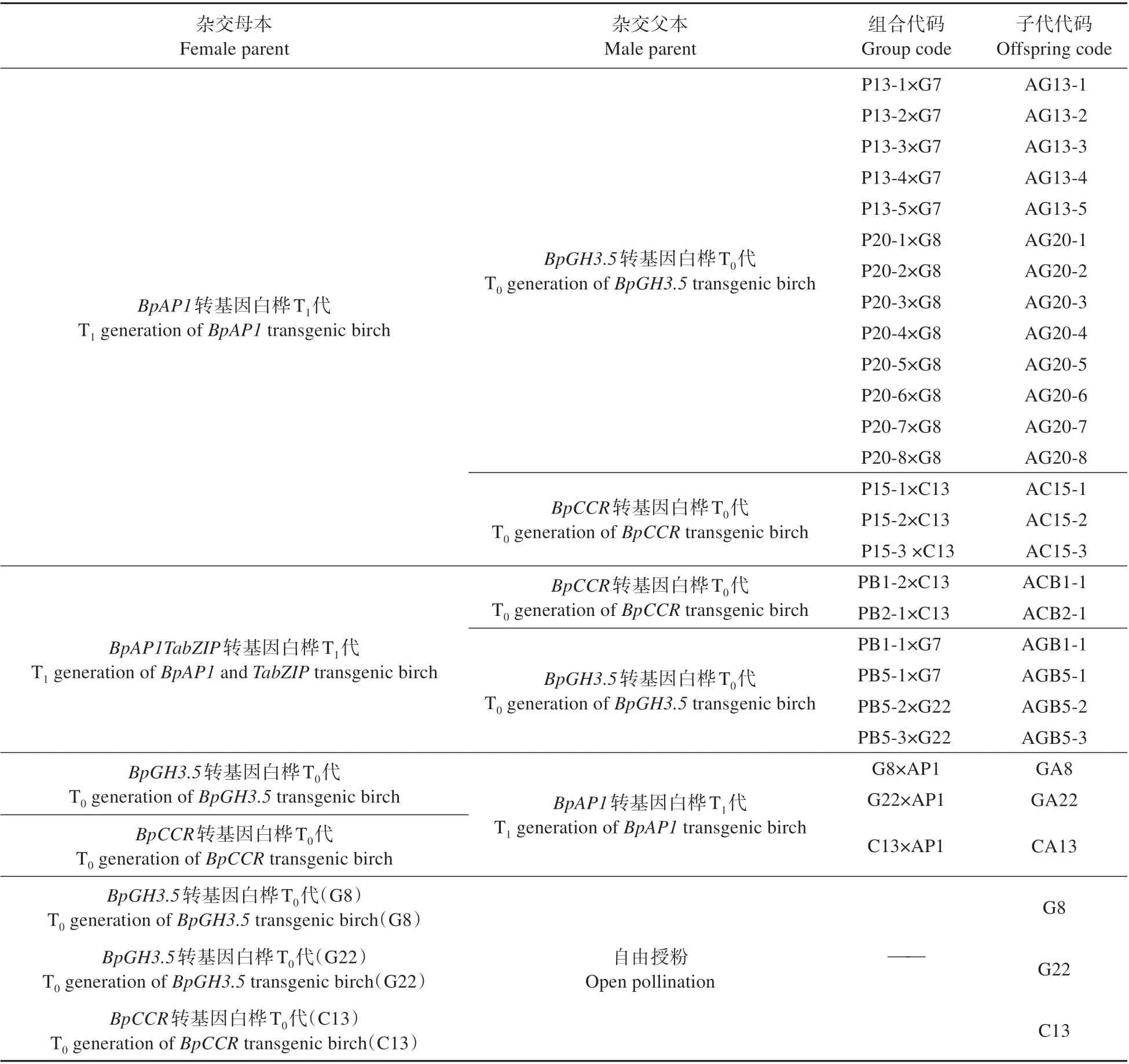
表1 参试白桦家系代码Table 1 Tested white birch family code
1.3.2 PCR检测
以白桦总DNA 为模板,对BpGH3.5、BpCCR 转基因杂交亲本进行PCR 扩增检测,设置中间质粒载体pGWB2 为阳性对照,非转基因白桦(WT)为阴性对照,同时设置空白水对照。分别根据Bp⁃GH3.5、BpCCR 两端序列设计引物(见表2)。PCR反 应 体 系 如 下:10×Taq buffer 2 µL,dNTP(2.5µmol·L-1)1.6 µL,上下游引物(10 µmol·L-1)各1µL,MgCl2(2.5 µmol·L-1)1.2 µL,DNA 1 µL,Taq 酶0.5 µL,最后用H2O 补齐至20 µL。PCR 扩增程序为:94℃预变性5 min,94℃变性30 s,60℃退火30 s,72℃延伸1 min,30 个循环,72℃延伸10 min。扩增产物用1.0%的琼脂糖凝胶进行检测。
2 结果与分析
2.1 BpGH3.5、BpCCR 转基因杂交亲本的PCR检测
以BpGH3.5、BpCCR 转基因白桦DNA 为模板,对待测株系进行PCR 检测,结果显示:4 株亲本植株G7、G8、G22、C13均检测到目的基因(见图1)。

表2 PCR引物序列Table 2 Primer sequences for PCR assays
2.2 不同杂交子代种子活力性状比较
2.2.1 不同杂交子代种子活力性状差异显著性分析
通过对各参试家系种子的千粒质量、发芽率、发芽势等指标进行方差分析,结果显示:各活力性状在不同子代家系间的差异均达到了极显著水平(P<0.01)(见表3)。其中除种子千粒质量和平均发芽时间的变异系数较小外,发芽率、发芽势、发芽指数等性状的变异系数均在40.00%以上,这表明种子发芽率、发芽势等性状在各家系间存在着较大的差异,因此,针对各子代家系进行选优意义重大。遗传力分析结果显示:种子千粒质量、发芽率、发芽指数等性状的家系遗传力均在95.00%以上,表明种子活力性状主要是由遗传因素控制的。
2.2.2 不同杂交子代种子活力性状的多重比较分析
在种子活力性状在各子代家系间差异达到极显著水平的基础上,对28个子代家系的种子千粒质量、发芽率、发芽势等性状进行多重比较分析,进一步了解各性状在家系间的关系。数据显示:白桦种子千粒质量较大的家系,除平均发芽时间外,其发芽率、发芽势、发芽指数等种子活力指数也相对较高,家系AG20-6的千粒质量高达381.50 mg,其发芽率、发芽势也分别达到54.02%、7.47%,在28个家系中排名靠前。发芽率是衡量种子品质好坏的一个重要指标,28个参试家系种子发芽率的多重比较分析结果显示(见表4):家系AG20-6、CA13、AG20-8和AG20-7的发芽率排名靠前,均在45%以上,分别为54.02%、46.09%、45.96%和45.94%,其中AG20-6最高。这4个家系的千粒质量、发芽势和发芽指数,除AG20-8的发芽势较低以外,其他排名也均靠前。
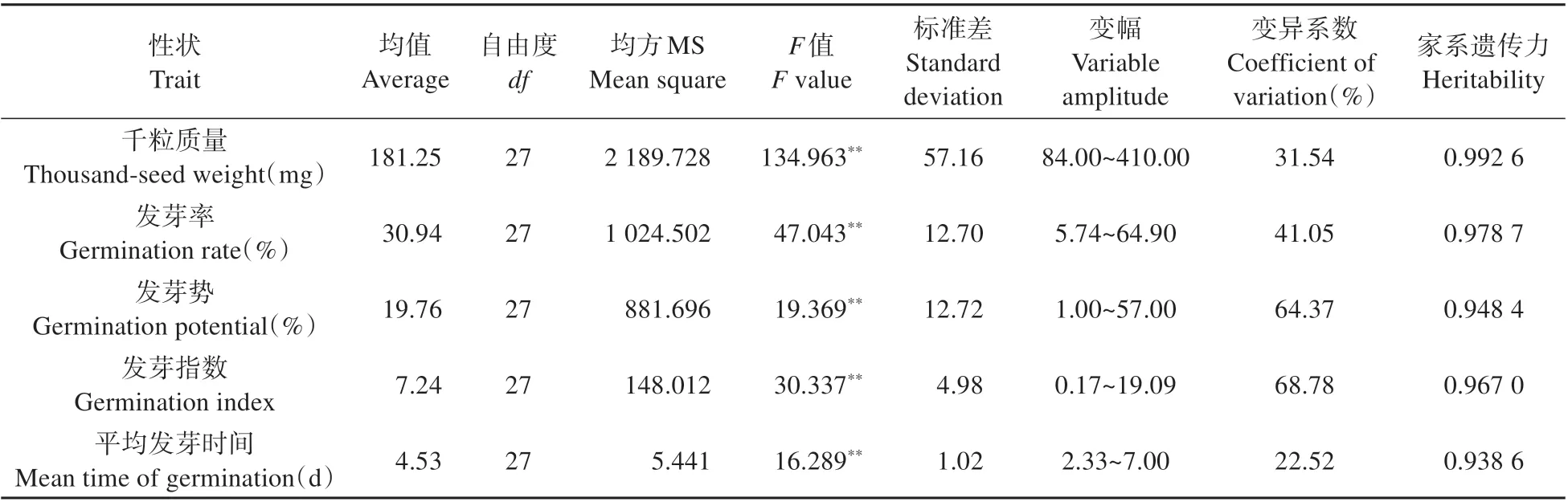
表3 不同杂交子代家系种子活力性状的方差分析表Table 3 Variance analysis among different hybrid offspring families on seed vigor index
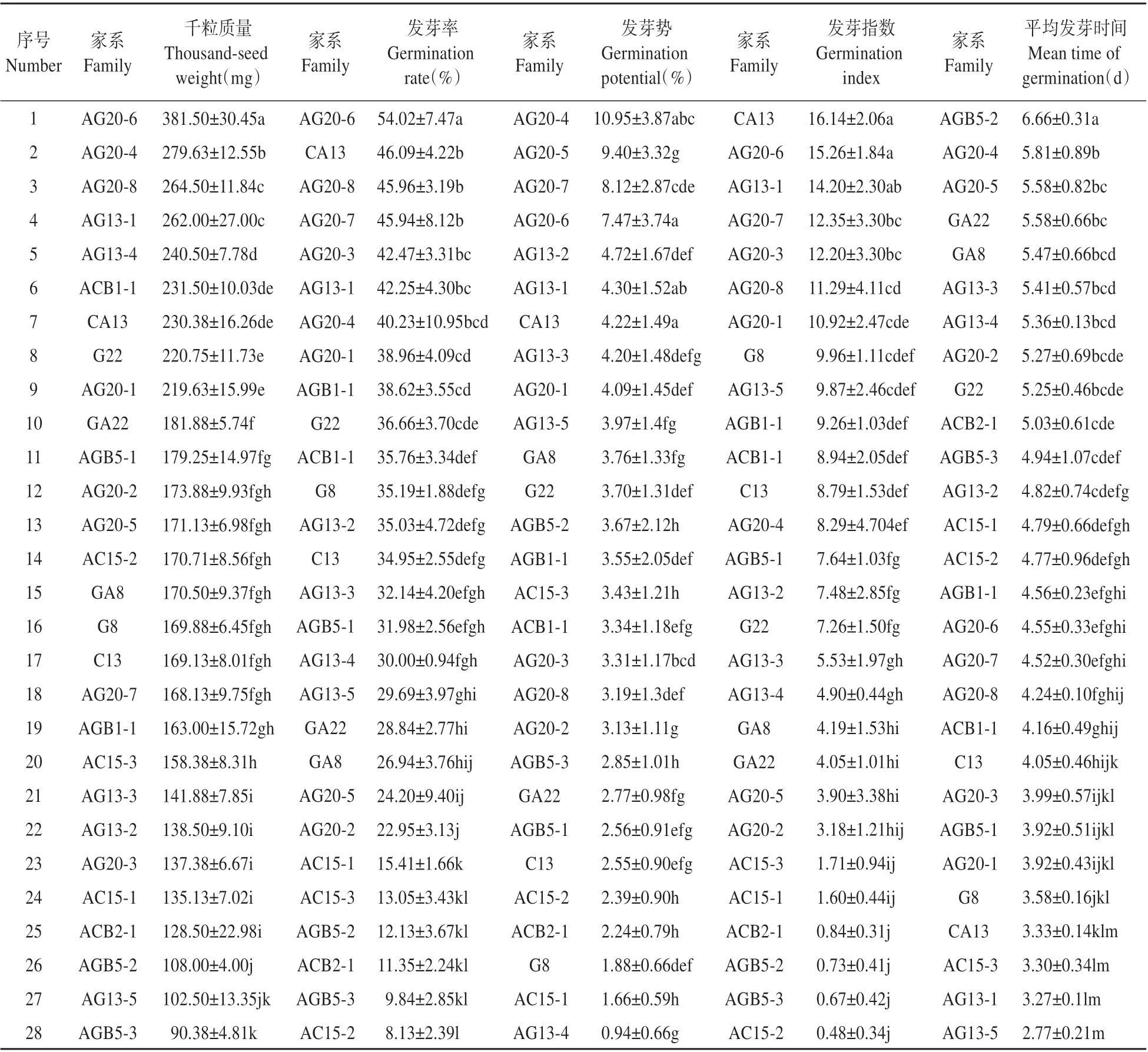
表4 不同杂交子代家系种子千粒质量、发芽率、发芽势、发芽指数、平均发芽时间的多重比较Table 4 Multiple comparison thousand-seed weigh,germination rate,germination potential,germination rate and mean time of germination among different families
2.2.3 不同杂交子代家系种子活力性状相关性分析
在对各性状进行多重比较分析的过程中发现,除平均发芽时间外,千粒质量与发芽率、发芽势、发芽值等性状在排序过程中表现出相对的一致性,因此针对各性状进行相关性分析(见表5),结果显示:千粒质量与发芽率、发芽势、发芽指数间 的 相 关 系 数 分 别 为0.611、0.603、0.528(P<0.01),其他性状间的相关系数均大于0.800,说明参试家系的种子活力指标之间具有极其显著的相关性。
2.3 外源基因在杂交子代中的传递规律
选取4 个杂交子代家系,进行叶片总DNA 的提取,采用PCR 扩增技术检测不同株系中目的基因(BpGH3.5、BpAP1、TabZIP、BpCCR)在杂交T2代中的分离情况,然后统计各基因型的株数及比例,利用卡方检测法对基因型的比例进行验证,分析外源目的基因插入位点数及在T2代中的传递规律。
由表6可见,目的基因能够通过一定的方式遗传给子代且不同转基因株系的子代外源基因检出率不同。通过对AG20-1、AGB5-1、AG20-4、ACB2-1这4 个家系进行PCR 检测,由于每个家系仅检测了12~31株幼苗,其中AG20-1、AGB5-1、AG20-4分别为24、31 和26 株,ACB2-1 更是显著小于30 株,PCR 检测结果的性状分离比例与理论性状分离比例可能会存在较大的偏差甚至不具有统计学意义,因此对4 个家系的检测结果进行适应性检验。根据AG20-1、AGB5-1、AG20-4、ACB2-1 群体中目的基因组合及分离的4种基因型株数,推测其比例为1∶1∶1∶1,通过卡方检验,其值分别为12.33、9.52、19.54、4.67,前三者均显著大于X20.05(3)(7.82),后者小于X20.05(3)(7.82)。其中由于ACB2-1 的检测样本数仅为12株,其检验误差较大,因此不能得出其目的基因能够按照孟德尔遗传规律进行传递的结论。对AGB5-1 和ACB2-1 家系子代进行PCR 的结果显示,AGB5-1 子代群体中检测出BpAP1、Bp⁃GH3.5,ACB2-1 子 代 群 体 中 检 测 出TabZIP、BpAP1,即未获得3个目的基因聚合的子代个体。
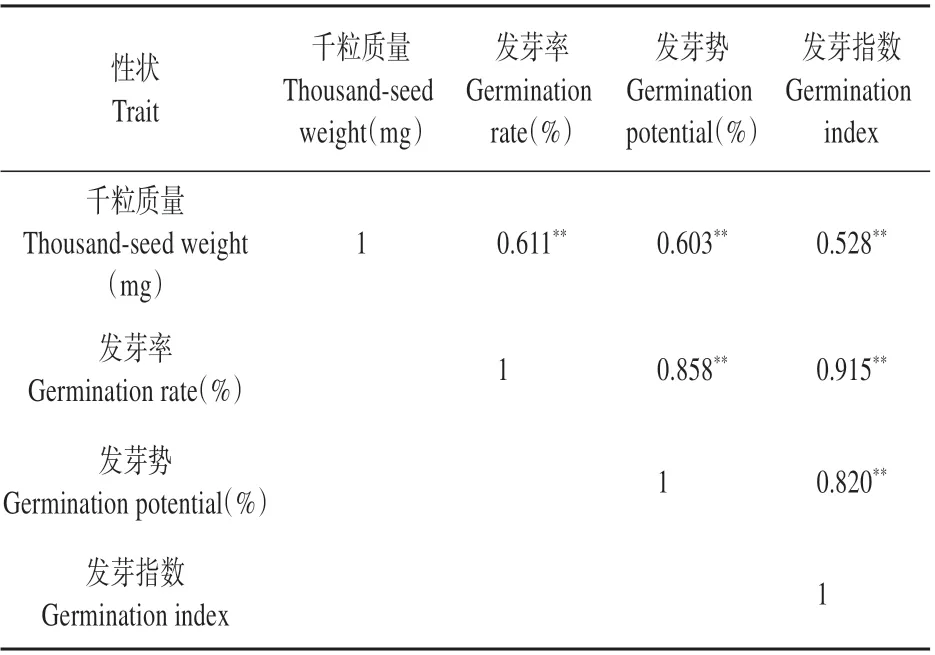
表5 不同杂交子代家系种子活力性状的相关分析Table 5 Correlation analysis among seed vigor charac‐ter of different families
2.4 杂交子代生长性状分析
对参试家系的苗高、苗高生长量、苗高相对生长量进行方差分析(见表7)研究表明:各性状在不同杂交子代家系间均达到极显著差异(P<0.01)。各性状的变异系数均在36.77%~60.00%,表明不同杂交子代苗高、苗高生长量、苗高相对生长量的整齐性一般。各性状的家系遗传力均在在0.85 以上,其中苗高和苗高生长量家系遗传力均为0.91,说明苗高、苗高生长量、苗高相对生长量主要是受遗传因素控制。
各性状在家系间差异达到极显著水平的基础上,对参试家系的苗高、苗高生长量等性状进行多重比较分析(见表8),进一步了解各性状在家系间的关系。结果表明:对苗高、苗高生长量、苗高相对生长量进行由大到小的排序,结果发现:G22、GA8 和GA22 的苗高最高,分别为52.30、51.03 和49.55 cm;这3 个家系的苗高生长量的排名也靠前,分别为48.08、44.46 和45.18 cm;相对生长量除GA8 低一些,其他两个家系的相对生长量仍然很高,为12.58 和11.93 cm。这说明家系间的苗高和苗高生长量表现出一致性,即苗高排名靠前的家系,其苗高生长量排名也靠前。苗高相对生长量与苗高和苗高生长量的一致性相对较差。
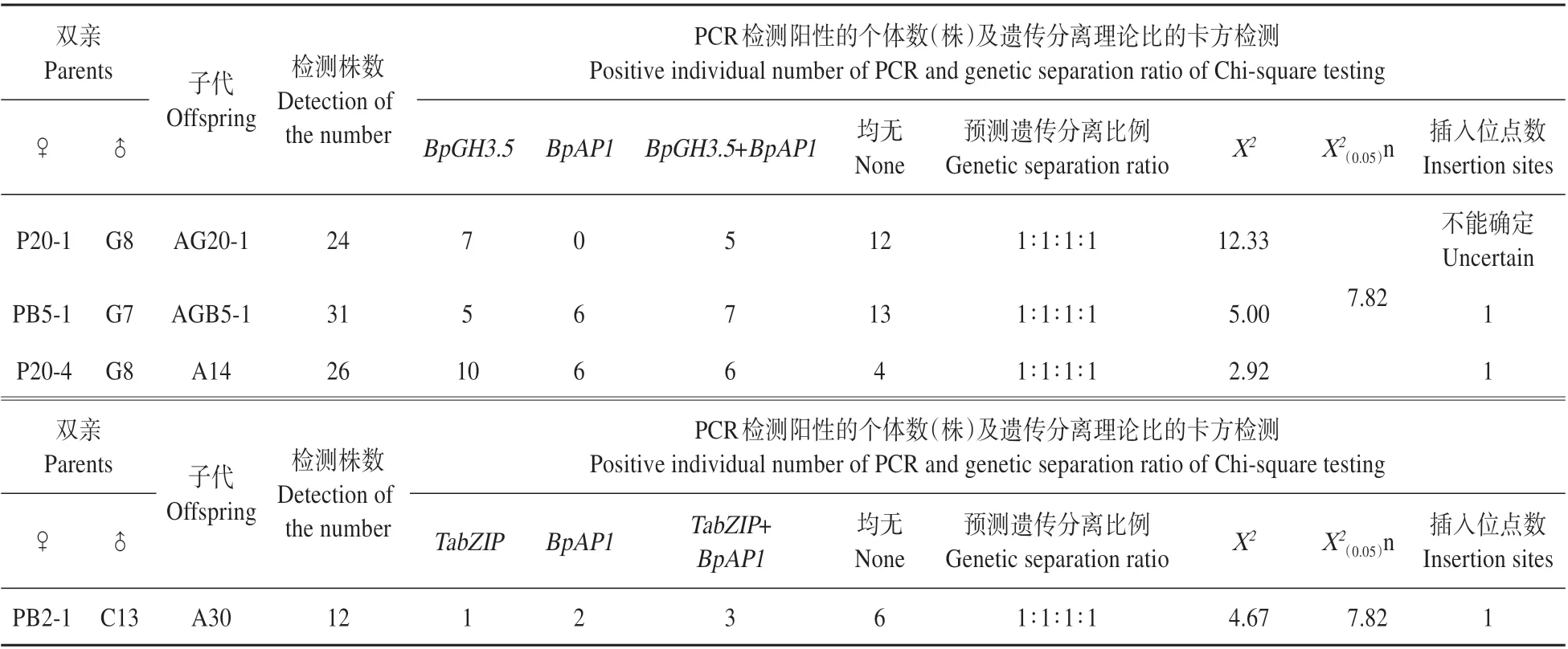
表6 外源基因在子代中的PCR鉴定及χ2检测Table 6 Detection of the foreign genes in offspring and Chi-square testing
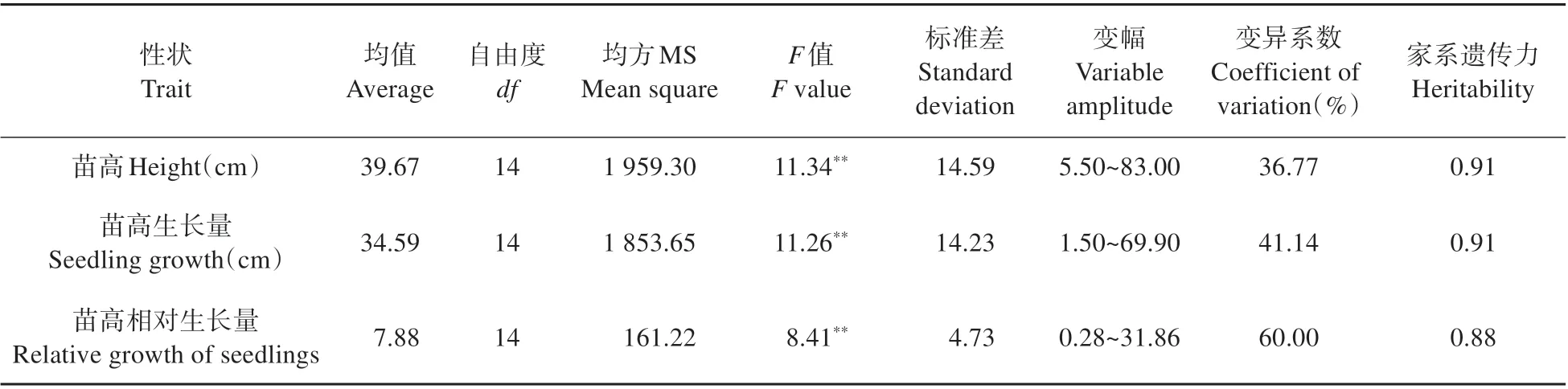
表7 不同杂交子代苗高、苗高生长量、苗高相对生长量的方差分析Table 7 Variance analysis on seedling height,seedling growth,relative growth of seedlings among different families

表8 不同杂交子代苗高、苗高生长量、苗高相对生长量多重比较Table 8 Multiple comparison seedling height,seedling growth,relative growth of seedlings among different families
3 讨论
林木转基因的目的是按照人们预期的育种目标培育良种[20]。单个基因的优良特性往往不能满足生产生活的需要。例如:抗病育种中,由于病菌的变异等因素,利用单基因的抗性进行防治就存在较大的风险,所以利用不同抗性基因的协同作用并实现聚合育种是当今学者研究的热点[21]。随着现代分子技术的飞速发展,利用基因工程等方法已在多种动植物上实现多基因的聚合育种。基因工程等分子育种手段能够快速获得多基因聚合的个体,而林木转基因体系较难建立且林木生长周期长,所以以缩短童期的BpAP1 转基因白桦为载体,采用常规育种方法研究林木转基因植株间的聚合育种是一个有效的途径。
本研究进行转基因植株间的杂交,测定了杂交子代千粒质量、发芽率等活力指标,结果发现种子活力指标间均呈现出极显著的正相关关系,这与陈素素[18]、王朔[22]等研究结果一致,千粒质量与发芽率、发芽势等指标呈极其显著的正相关关系,认为千粒质量是种子活力的基础。此外,本研究发现目的基因的随机插入降低了白桦杂交子代种子活力,其AG20-1、AGB5-1、AG20-4、ACB2-1 的发芽率分别为38.96%、31.98%、40.23%和11.35%。目的基因的随机插入对杂交子代活力的影响与正反交有着密切的联系,除CA13 外,GA18、GA22 的发芽率仅为26.94%、28.84%,表明雌配子的稳定遗传能力较雄配子强,且目的基因BpCCR 对白桦种子活力的影响较目的基因BpGH3.5 小。采用PCR 扩增法对杂交T2代的外源基因进行检测,结果发现插入母本基因组中BpAP1 基因的检出率分别为20.83%和46.15%,而插入父本基因组中Bp⁃GH3.5 基因的检出率分别为50.00%和61.54%,显然杂交T2代群体中插入父本基因组的目的基因检出率较高,其雌雄配子的传递效率分别为30.00%和50.00%左右。陈素素等[18]有关转基因白桦T1代的研究表明,雌配子的传递效率要高于雄配子,周静等[24]认为由于外源基因的插入,植物花粉的发芽力、受精能力等均受到了不利影响,因而雌配子的传递效率较雄配子高。本研究与前人的研究结果偏差较大,认为目的基因通过转基因植株间杂交而进行传递,在T2杂交子代中,其种子活力进一步降低,同时由于1年生BpAP1转基因白桦T1代和多年生BpGH3.5、BpCCR转基因白桦的插入位点不能确定,导致其杂交T2代群体中插入位点不能确定,目的基因在T2代的遗传规律是否符合孟德尔遗传规律有待进一步研究证明。有研究表明,目的基因在转基因植株子代的传递规律呈多样性,通常转基因植株中有10%~50%的频率不符合孟德尔遗传规律[23~26],因此有关转基因白桦目的基因在子代群体中传递规律需要进一步进行验证。
外源基因的整合对发芽率等性状的影响效果不 显 著[18],结 合PCR 检 测 结 果,认 为 以1 年 生BpAP1 转基因白桦T1代为母本,多年生转基因白桦为父本的杂交过程中,T2代种子的发芽率大幅度降低,初步说明外源基因的再次传递降低种子活力。本研究未获得3目的基因聚合的T2代个体,在对T2代群体长期监测的基础上同时结合PCR 检测结果,许多实现基因聚合的子代个体往往是生长状况较差的个体,且许多幼苗个体在成苗后的一定时期便发生枯死现象,研究认为是有目的基因的聚合,多基因的叠加效应对白桦转基因杂交子代个体的生长发育产生了不利的影响,并可能导致致死现象,因此未能获得3目的基因聚合的T2代个体。
常规育种方法具有简单、易操作的特点。研究表明利用常规育种方法开展转基因白桦间的聚合育种研究是具有可行性的。转基因白桦间杂交后,目的基因在其杂交子代的传递规律,多个目的基因聚合后对白桦子代个体生长产生的影响以及表达调控机制等都是我们关注的重点,后续将利用缩短童期的BpAP1 转基因白桦作为优良载体,采用常规育种方法并结合分子标记、二代测序等分子技术,进一步探讨转基因白桦间杂交后,目的基因在子代群体的遗传规律。
猜你喜欢
今日农业(2022年6期)2022-07-05
科学导报(2020年8期)2020-03-12
安徽农学通报(2020年1期)2020-02-29
山东农业科学(2019年11期)2019-12-24
文苑(2019年20期)2019-11-20
文苑·经典美文(2019年10期)2019-10-16
现代农业科技(2019年1期)2019-07-11
现代农业科技(2017年24期)2018-01-22
华人时刊(2016年16期)2016-04-05
现代园艺(2016年2期)2016-03-15
