
猪繁殖与呼吸综合征病毒GP5蛋白胞外区的串联表达及免疫原性分析
2021-04-20 01:21金前跃王寅彪冯丽丽李华玮郭振华邢广旭张改平
现代牧业 2021年1期
金前跃,王寅彪,冯丽丽,李华玮,郭振华,邢广旭,张改平,4
(1.河南省农业科学院,动物免疫学重点实验室,河南郑州 450002;2.新乡医学院,公共卫生学院,河南新乡 453003;3.河南省农业科学院,农业经济与信息研究所,河南郑州 450002;4.河南农业大学,动物医学院,河南郑州 450002;5.河南牧业经济学院,动物医药学院,河南郑州 450046)
猪繁殖与呼吸综合征(Porcine reproductive and respiratory syndrome,PRRS)以母猪繁殖障碍及猪群呼吸困难为主要特征,在世界范围内流行并严重制约养猪产业的健康发展[1]。20世纪90年代,科学家们从临床患病猪身上分离到病毒,确定了引起该病的主要病原体为PRRSV[2, 3]。PRRSV为有囊膜RNA病毒,属动脉炎病毒科(Arteriviridae),动脉炎病毒属(Arterivirus),根据分子生物学特征分为欧洲型(I型)和美洲型(II型)[4-6]。1996年,我国首次分离到该病毒,研究发现为美洲型[7]。2006年,新型的毒力更强的美洲型PRRSV在我国十几个省广泛流行[8]。尽管我国报道的PRRSV大多属于美洲型,但是新的欧洲型PRRSV也开始在我国流行[9]。因此,有必要建立PRRSV血清抗体检测方法以用于该病的流行病学调查;而且,为了对PRRSV的疫苗效力及PRRS疾病进程进行判断和评价,建立PRRSV中和抗体的检测方法亦是十分必要。
GP5蛋白是PRRSV的主要结构蛋白之一,在病毒中和方面起重要作用[10, 11]。该蛋白为跨膜蛋白,其胞外区与M蛋白胞外区构成异源二聚体[12-14]。GP5-ecto与M蛋白胞外区形成的异源二聚体能够介导PRRSV入侵宿主细胞,其中GP5蛋白诱导产生的中和抗体的功能是提供免疫保护,M蛋白则诱导产生强烈的细胞免疫,因此,GP5蛋白是良好的亚单位疫苗设计靶标[15-19]。Plagemann等利用重叠多肽合成技术证实PRRSV主要中和表位在GP5-ecto的中间部位[20, 21],故GP5-ecto成为建立检测PRRSV中和抗体的首选靶标。
本研究以GP5-ecto基因为对象,通过基因合成技术串联合成该基因片段,利用大肠埃希氏菌E.coliBL21(DE3)对该片段进行表达,并纯化获得具有良好免疫原性的蛋白。本研究为PRRSV中和抗体ELISA检测方法的建立奠定了基础。
1 材料和方法
1.1 材料
1.1.1 病毒、载体和菌株
PRRSV河南株HN07-1(GenBank No. FJ147205.1)、MARC-145细胞、原核表达载体pET28a、大肠埃希氏菌感受态细胞JM109和E.coliBL21(DE3)、PRRSV阳性血清及阴性血清由河南省农业科学院动物免疫学重点实验室保存。
1.1.2 酶及主要试剂
Premix ExTaq DNA聚合酶、DL 2000 DNA Marker、限制性内切酶购自TAKARA;DNA连接试剂盒、DNA纯化回收试剂盒、质粒提取试剂盒购自TIANGEN;HRP标记二抗、His单抗购自北京博奥森;IPTG购自华美公司;DMEM培养基及新生胎牛血清购自SOLARBIO;弗氏完全及不完全佐剂购自GIBCO。
1.2 方法
1.2.1 PRRSV-GP5胞外区基因的串联合成
本实验将PRRSV HN07-1株GP5蛋白的胞外区在基因水平上进行串联连接,引入BamH I和XhoI的酶切位点。GP5-ecto序列交由生工生物工程(上海)有限公司合成并克隆到载体pBluescript II SK+上,以下为对应的氨基酸序列:SNNSSSHIQLIYNLTLCELNGTDWLTQKFDWAHHHHHHH
HSNNSSSHIQLIYNLTLCELNGTDWLTQKFDWA。
1.2.2 重组原核表达质粒pET28a-GP5-ecto的构建及鉴定
使用BamH I和XhoI进行双酶切将GP5-ecto序列从pBluescript II SK+载体上切除后与同样经BamH I和XhoI双酶切的pET28a载体用DNA连接试剂盒进行连接。连接产物pET28a-GP5-ecto转化大肠埃希氏菌感受态细胞JM109,涂布Kan+抗性的LB固体平板,置于37 ℃培养并挑选单克隆菌落。鉴定后的阳性重组质粒转化大肠埃希氏菌感受态细胞E.coliBL21(DE3)并挑取单克隆菌落用以诱导重组蛋白GP5-ecto的表达。
1.2.3 重组蛋白GP5-ecto的表达、纯化及免疫原性鉴定
将含重组质粒pET28a-GP5-ecto的阳性单克隆菌落在液体LB培养基中培养至菌液OD600值为0.6~1时,以终浓度为0.5 mM的IPTG,在37 ℃条件下诱导4 h。通过SDS-PAGE鉴定GP5-ecto蛋白的诱导表达情况,并用Western blot鉴定GP5-ecto蛋白与PRRSV阳性猪血清的反应。使用Ni-NTA亲和层析对GP5-ecto蛋白进行纯化并采用尿素浓度梯度复性后,使用ELISA检测方法鉴定其与PRRSV阳性猪血清的反应情况,以此来判定GP5-ecto蛋白的免疫原性。
ELISA检测方法如下:将纯化蛋白以不同浓度在4 ℃进行过夜包被;PBST清洗3次,5%脱脂奶于37 ℃封闭1 h;PBST清洗3次,加入1∶400稀释的阳性及阴性血清,37 ℃下孵育0.5 h;PBST清洗3次,加入1∶1000倍稀释的兔抗猪HRP-IgG,37 ℃孵育0.5 h;PBST清洗3次,使用TMB显色液进行显色,酶标仪进行检测。
1.2.4 重组蛋白GP5-ecto的BALB/c小鼠免疫及IFA试验
使用纯化后的GP5-ecto蛋白免疫SPF级6~8周龄雌性BALB/c小鼠,共免疫3只,进行3次免疫。首免使用弗氏完全佐剂与GP5-ecto蛋白乳化,采用背部皮下多点注射的方法每只注射50 ng蛋白。7天后,进行二免,使用弗氏不完全佐剂与GP5-ecto蛋白乳化,采用同上剂量进行免疫。14天后,进行三免(方法同二免)。三免10天后采集小鼠血清,使用IFA检测GP5-ecto蛋白免疫后的小鼠血清是否具有与天然PRRSV病毒蛋白反应的能力。
IFA检测方法如下:将MARC-145细胞在96孔板中培养至80%左右,接种PRRSV HN07-1病毒;感染48 h后,弃去上清,用预冷PBST清洗3次,含0.3%H2O2的预冷甲醇溶液固定15 min;弃固定液,PBST清洗3次,5%脱脂奶37 ℃封闭1 h;PBST清洗3次,加入1∶100倍稀释的免疫血清和PRRSV阳性及阴性血清,37 ℃孵育1 h;PBST清洗3次,分别加入1∶100倍稀释的羊抗鼠FITC-IgG或兔抗猪FITC-IgG,37 ℃孵育1 h;PBST清洗3次,荧光显微镜观察结果。
2 结果与分析
2.1 重组原核表达质粒pET28a-GP5-ecto的鉴定
重组质粒pET28a-GP5-ecto经NdeI和XhoI、BamHI和XhoI两组双酶切鉴定后均出现特异目的条带,表明重组质粒连接成功(图1)。
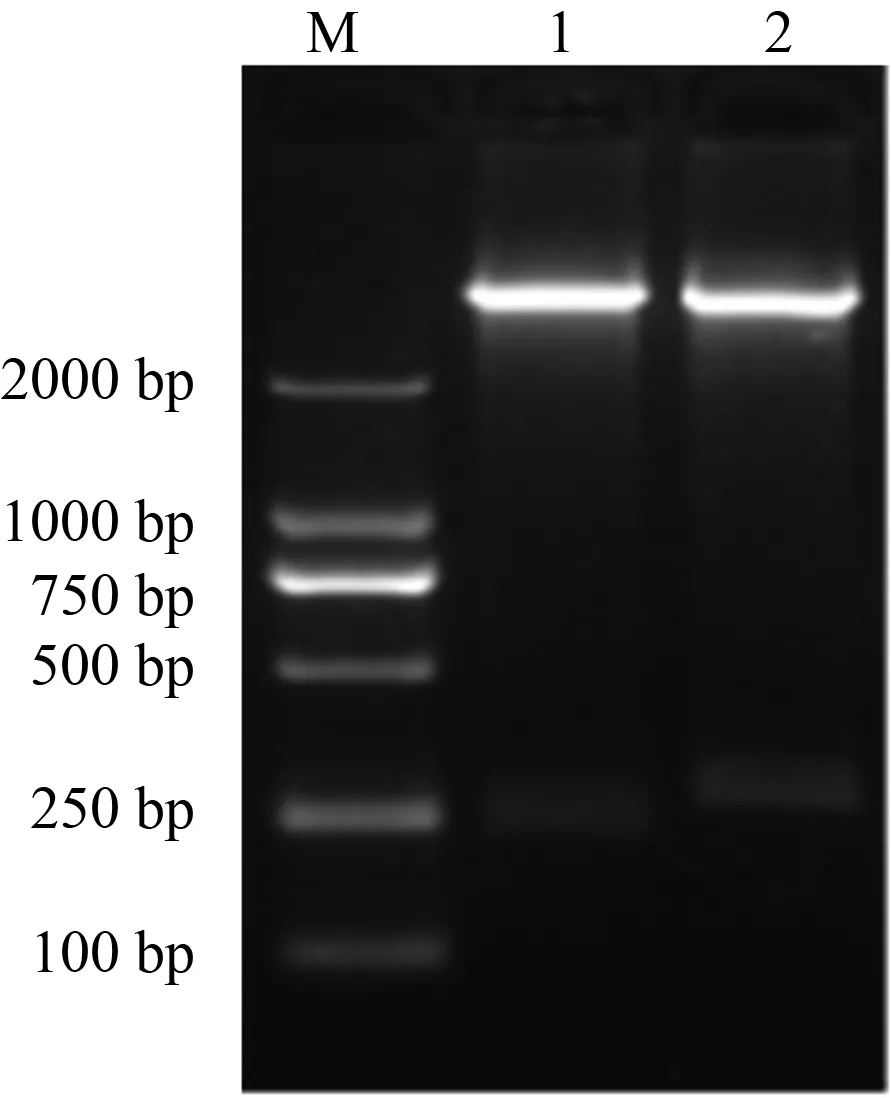
M. DL 2000 (Marker); 1. BamH I和Xho I双酶切;2. Nde I和Xho I双酶切图1 重组原核表达质粒pET28a-GP5-ecto的鉴定
2.2 重组蛋白GP5-ecto的表达、纯化及Western blot鉴定
SDS-PAGE对重组蛋白GP5-ecto的表达情况进行鉴定后发现在12 kDa左右(理论计算值为12.3 kDa)有一条新出现的蛋白条带。而且在37 ℃培养至OD600值为0.6~1.0时,以终浓度为0.5 mM的IPTG诱导4 h后,重组GP5-ecto蛋白实现了高效表达。Ni-NTA亲和层析纯化后,采用不同浓度梯度的尿素进行复性。Western blot表明GP5-ecto蛋白能与PRRSV HN07-1阳性猪血清发生特异性反应(图2)。
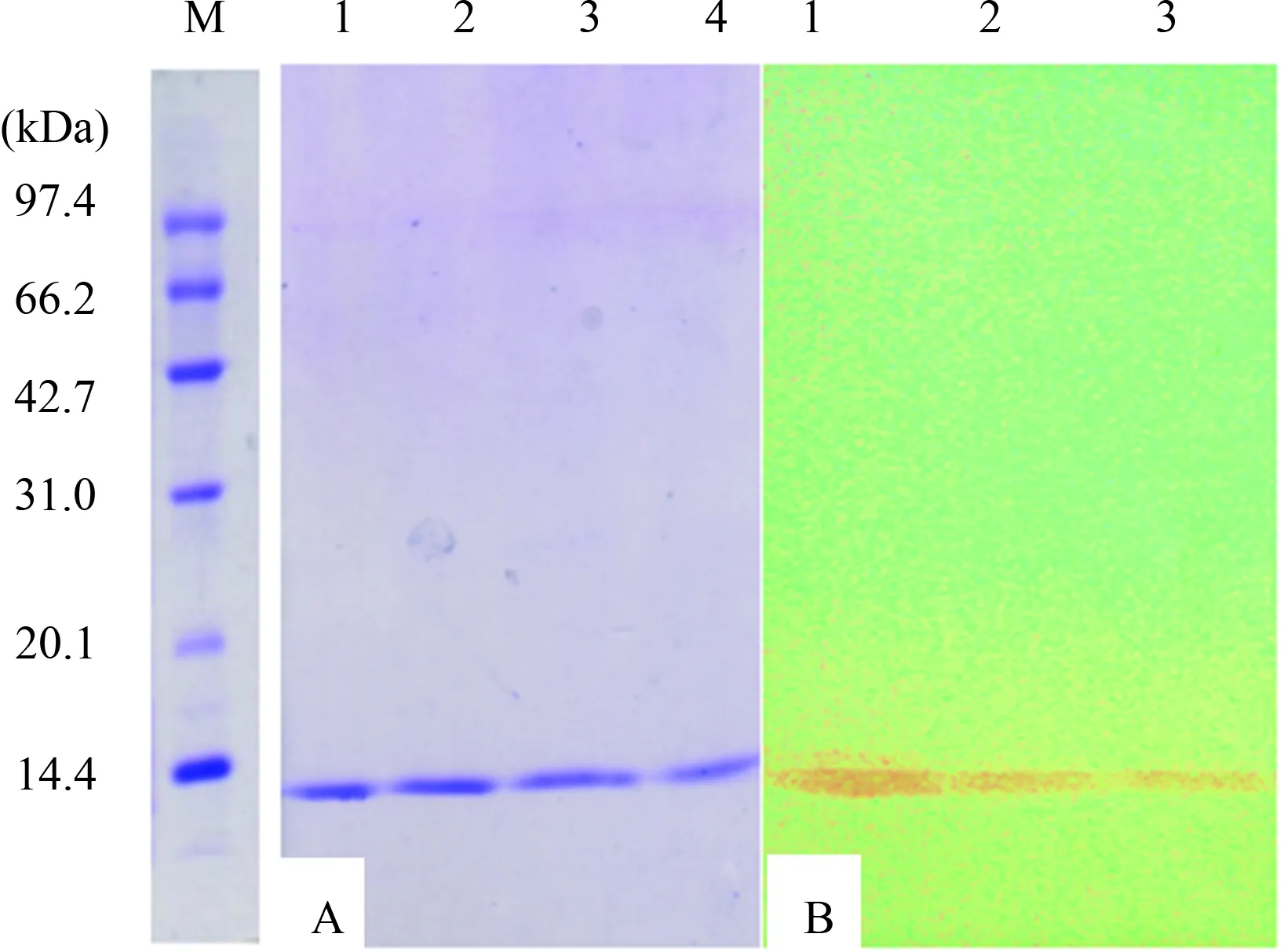
M. 低分子量蛋白质Marker; A. 1~4, SDS-PAGE鉴定GP5-ecto蛋白的表达; B. 1~3, Western blot鉴定纯化后的GP5-ecto图2 重组蛋白GP5-ecto的纯化及Western blot鉴定
2.3 重组蛋白GP5-ecto的免疫原性鉴定
实验运用ELISA方法鉴定重组GP5-ecto蛋白的免疫原性,将不同浓度蛋白过夜包被后, 以PRRSV阳性血清作为一抗进行检测。结果显示:重组蛋白能与阳性血清发生强烈反应,最低包被浓度达0.125 μg/mL,说明该纯化蛋白免疫原性较好(图3)。
2.4 IFA
在重组GP5-ecto蛋白免疫BALB/c小鼠后,采集小鼠血清,以PRRSV HN07-1株的阳性血清作为阳性对照,采用IFA实验对小鼠血清与病毒的反应性进行鉴定。结果表明,免疫血清能与病毒发生反应,说明重组蛋白与病毒天然蛋白结构具有较高的相似性(图4)。
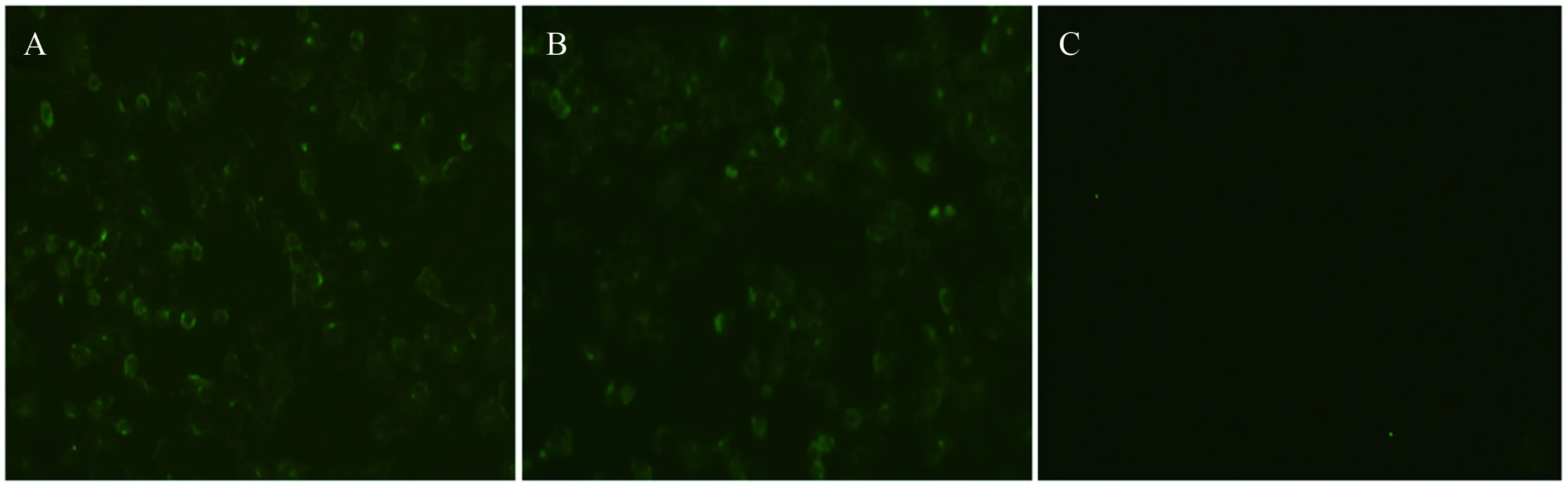
A. 免疫血清(1∶100); B.阳性对照(1∶100); C. 阴性对照(1∶100)图4 IFA实验结果
3 讨论与结论
我国现阶段存在传统美洲型PRRSV、高致病性美洲型PRRSV和欧洲型PRRSV同时流行的特点,面对如此严峻的疫病流行形势,急需开展疫病的长期监测,同时开发相应的诊断技术及产品。作为PRRSV主要的囊膜蛋白,GP5在PRRSV的诊断、血清流行病学监测及疫苗效力评价等方面都具有重要作用。
PRRSV感染后,康复期的猪血清中的中和抗体主要是针对GP5的蛋白[11]。GP5蛋白的单克隆抗体在病毒中和方面比GP4蛋白的单克隆抗体更有效[18]。Plagemann等将PRRSV的主要中和表位定位在GP5-ecto的中间部位[20]。Ostrowski等报道:GP5-ecto区域包含有中和性的B表位以及非中和性的A表位,而且A表位是免疫优势表位,在一定程度上抑制了B表位及中和抗体的诱导产生[21]。所有这些研究都表明了GP5蛋白及GP5-ecto区域在诱导中和抗体产生方面发挥重要作用,同时也为使用GP5-ecto区域作为靶标检测PRRSV中和抗体提供了重要依据。
GP5蛋白在重组卡介苗(BCG)、转基因植物及宿主动物等系统中均获得表达[15, 22, 23]。但考虑到成本及表达水平,大肠埃希氏菌原核表达系统是较好的选择。而GP5蛋白的跨膜区具有强疏水性,能够在一定程度上影响其表达[24]。课题组曾尝试使用大肠埃希氏菌原核表达GP5蛋白(仅缺失N端信号肽),但发现重组pET28a-GP5和pET32a-GP5在大肠埃希氏菌E.coliBL21(DE3)及E.coliRosetta细胞中均不表达,也从侧面证明了GP5蛋白跨膜区的存在能够影响其在大肠埃希氏菌中的表达。鉴于上述情况,我们在本研究中利用基因合成技术串联合成了GP5蛋白的胞外区,并利用大肠埃希氏菌E.coliBL21(DE3)表达系统成功表达了GP5-ecto蛋白,ELISA和IFA证实GP5-ecto蛋白具有良好免疫原性。本研究获得的GP5-ecto蛋白为检测PRRSV血清中和抗体的ELISA方法的建立提供了材料。
猜你喜欢
——一道江苏高考题的奥秘解读和拓展
中学生物学(2022年7期)2022-09-07
成都医学院学报(2022年4期)2022-08-19
文萃报·周五版(2021年34期)2021-09-13
医学理论与实践(2021年15期)2021-08-05
现代临床医学(2021年4期)2021-07-31
昆明医科大学学报(2021年2期)2021-03-29
爱你·健康读本(2020年8期)2020-08-23
爱你(2020年22期)2020-08-18
三农资讯半月报(2020年11期)2020-06-21
江苏农业学报(2019年1期)2019-09-10
