
下丘脑-垂体-肾上腺皮质轴应激反应的调控
2021-04-26 13:59冯晓丽任晓峰
探索科学(学术版) 2021年1期
冯晓丽 任晓峰
1.昆明医科大学 云南 昆明 650500
2.中国科学院动物模型与人类疾病机理重点实验室&中国科学院昆明动物研究所 云南 昆明 650223
3.中国科学院大学昆明生命科学学院 云南 昆明 650204
导言
应激反应是所有生物对紧张性事件的适应性反应,指机体在不良的外界环境因素(各种应激源)的应激下引起下丘脑、垂体和肾上腺皮质的非特异性神经、体液反应,使机体得以克服刺激的危害性,适应环境和维持机体内环境的平衡[1]。应激反应最主要的内分泌系统的变化是HPA轴的激活[2]。
HPA轴是下丘脑-垂体-肾上腺皮质轴的简称。下丘脑位于丘脑下方,紧靠第三脑室壁,每侧下丘脑分3个区:外侧区、内侧区和室周区。HPA轴中的“下丘脑”实际上特指下丘脑室周区的小细胞性神经分泌细胞。垂体位于脑基底部的下方,分为前叶和后叶。HPA轴中的“垂体”特指垂体前叶,即腺垂体,它的细胞合成和分泌多种激素,调节全身其他腺体的分泌。肾上腺位于肾脏的正上方,由外壳的肾上腺皮质和中心的肾上腺髓质组成[3]。
下丘脑室周区的小细胞性神经分泌细胞控制垂体前叶,它们的轴突不延伸到垂体前叶,而是通过门脉循环与垂体前叶联系。下丘脑-垂体门脉循环是位于第三脑室底部的毛细血管网,这些小血管向下延伸,在垂体前叶形成分支[4]。
下文分别介绍了机体在正常应激状态下和慢性应激(或抑郁)状态下HPA轴的分泌特点、下丘脑室旁核对HPA轴的激活,海马和杏仁核对HPA轴活动的控制、内侧前额叶皮质对HPA轴活动的调节。
1 应激状态下HPA轴的分泌特点
当机体发生应激反应时,下丘脑室旁核相关神经元分泌CRH和血管升压素增多,二者协同分泌。这两种激素进入门脉循环,并经血流向下运输,直到与垂体前叶细胞表面的特异性受体结合。这些受体激活后,使垂体细胞分泌ACTH。ACTH通过血液循环被运送到肾上腺皮质,与相应受体结合,激活受体,促使皮质类固醇激素的分泌,包括糖皮质激素,主要是皮质醇,盐皮质激素和少量性激素[4]。
糖皮质激素对于机体物质代谢的调节有重要作用,可以提高神经系统的兴奋性,在应激过程中参与提高机体耐受性。在下丘脑和垂体神经元及其他脑区广泛存在糖皮质激素的受体,在正常机体中,糖皮质激素大量分泌后,可通过血液的运输与这些受体结合,启动HPA轴本身的负反馈抑制,从而减少CRH的分泌,最终使糖皮质激素的浓度维持在一稳定水平。同时,ACTH的受体也存在于下丘脑CRH神经元上,起负反馈的作用,抑制CRH的过量分泌[3,4]。
在长期慢性应激的情况下,下丘脑持续处于激活状态,使得CRH过量分泌,导致糖皮质激素水平异常增高[5],地塞米松抑制不能(地塞米松是一种人工合成的肾上腺皮质激素,给正常个体注射地塞米松,会引发负反馈抑制作用,从而抑制HPA轴的分泌,但在抑郁患者当中这种负反馈机制受损,不能正常完成上述负反馈抑制)[6],后者说明经受长期应激的个体不能正常完成上述负反馈抑制。事实上,导致HPA轴过度活跃的因素常常来自生活中持续存在的应激,进而使得肾上腺把过多的皮质醇释放到血液中。
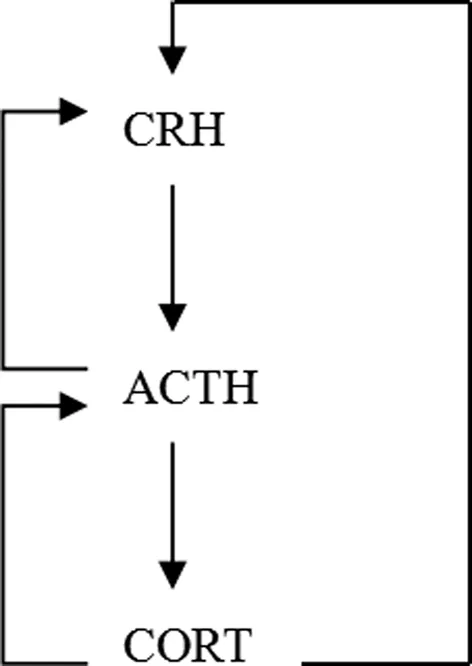
图1 HPA轴的分泌反应过程
2 下丘脑室旁核与HPA轴的激活
下丘脑的室旁核(PVN)是HPA轴活动的直接控制部位,下丘脑神经元可接受中枢和外周神经系统其他部位传来的神经信息,其神经分泌细胞分泌CRH控制ACTH的释放,室旁核的其它细胞调控自主神经系统。在实验动物脑内注射CRH可以引发明显的应激反应和抑郁症的典型症状,动物出现睡眠障碍、食欲下降和性行为减少[4]。有研究表明,通过腰椎穿刺发现,抑郁症病人脑脊液中CRH的含量高于其他精神病患者和正常人。如此一来,CRH就被认为是抑郁症及其主要行为反应的内分泌病理基础[5]。
动物实验及临床证据都表明,早期的不利经验与成年期发生抑郁的比例之间有相关性[7,8],母爱剥夺或早期不利经历都和脑内CRH神经元的持续的过度活动相关[3],所以Charles Nemeroff提出,CRH神经元持久的过度活动可能是早期不利经验容易导致成年发生抑郁的机制之一。他的研究小组还致力于研究一些新旧抗抑郁药物的作用,除了确定这些药物对单胺能神经元的作用之外,还要进一步确定它们的作用机制是否在于降低了CRH的分泌[3]。
同时,将抗抑郁药研究的焦点集中于CRH受体拮抗剂有着广泛的应用前景[5]。因为这类药物与CRH竞争垂体ACTH细胞表面的CRH受体,最终使HPA轴的糖皮质激素分泌水平降低,起到抗抑郁的效果。
3 海马、杏仁核对HPA轴活动的控制
(1)海马对HPA轴活动的控制。海马是HPA轴活动调节的高位中枢,在正常机体中,海马对HPA轴活动存在抑制性效应。兴奋海马引起实验动物如大鼠或者人的HPA轴活动降低;损害海马则使实验动物如大鼠和非人灵长类动物在应激反应时糖皮质激素分泌增加,并使PVN小细胞神经元表达CRH和AVPmRNA增加。这表明海马对HPA轴的活动有抑制影响,这种影响可能通过调节PVN的活动而产生[9]。
相对于其他应激相关脑区,糖皮质激素受体在海马的分布密度最大[6,9],在正常应激反应当中产生的大量糖皮质激素可以与之结合,激活受体,使海马对下丘脑室旁核神经元的抑制性输出起作用,从而减少ACTH和糖皮质激素的分泌[10]。
在抑郁个体中,海马的糖皮质激素受体功能存在缺陷。抑郁患者体内糖皮质激素水平过高,引起海马区域中含糖皮质激素受体的细胞数量减少,从而减低糖皮质激素与海马区域的相应受体的结合水平,使在海马水平的负反馈作用产生紊乱。
Carmine M.Pariante等研究者从抗抑郁药物的角度出发,着手研究糖皮质激素受体系统的变化对转基因抑郁小鼠(小鼠糖皮质激素受体功能缺陷)的影响,实验发现,在没有内源性的类固醇的情况下,抗抑郁药物可以诱导糖皮质激素受体向细胞核内转移,增强地塞米松诱导的受体转移,但是并未影响糖皮质激素受体蛋白质的表达[11]。另外,PW Gold和GP Chrousos的研究表明,抑郁病人的垂体水平的糖皮质激素负反馈是正常的[12],同时又鉴于抗抑郁剂能产生上述效应的时间与其纠正HPA系统活动的作用时间相一致[6,13],所以将海马区的糖皮质激素受体作为抗抑郁药的作用靶点具有一定的可行性[14]。
(2)杏仁核对HPA轴活动的控制。杏仁核是机体重要的情绪调控系统。在机体内,感觉信息进入杏仁核的基底外侧部,在那里进行处理和中继后,传送至杏仁核其他部位的神经元。与海马的作用不同,杏仁核对HPA轴的活动起兴奋性作用,Mason在非灵长类动物的研究证实了电刺激杏仁核激活HPA轴的活动[9]。
设计更为精细的动物研究表明,主要是CEA(中央核,为自主神经运动区)、MEA(内侧核,为自主神经运动区)和COA(嗅皮质,属于嗅系统)参与激活HPA轴的应激反应[9]。同时,杏仁核还可以激活脑干的去甲肾上腺素能核团(通过CRH激活脑干蓝斑系统,冲动经过延髓传递到脊髓的交感神经节前纤维,命令肾上腺髓质分泌儿茶酚胺,此即肾上腺髓质的应激反应),间接调节应激反应。
在正常的应激反应中,杏仁核分泌的CRH可以通过血液循环被运输到垂体,促使后者分泌ACTH,然后与肾上腺皮质的特异受体结合,释放皮质激素。
而在人类抑郁患者观察到左侧杏仁核代谢的活跃程度增高,并且抑郁程度越严重,脑脊液皮质醇浓度越高,左侧杏仁核代谢水平越高。抗抑郁药则能降低这种病理性代谢活跃程度[10]。以上这些都说明杏仁核对患者脑内皮质醇的增多起促进作用,而后者同样可以增强杏仁核的活动性。Shepard通过给正常大鼠注射皮质酮,使其杏仁体中央核(除了下丘脑室旁核,脑内另一生产CRH的主要区域)的CRHmRNA水平上升[15]。
事实上,海马和杏仁核没有纤维直接投射到下丘脑室旁核神经元,它们对HPA轴活动的控制都可能是通过下丘脑的局部GABA能神经元回路而发挥作用的[16]。
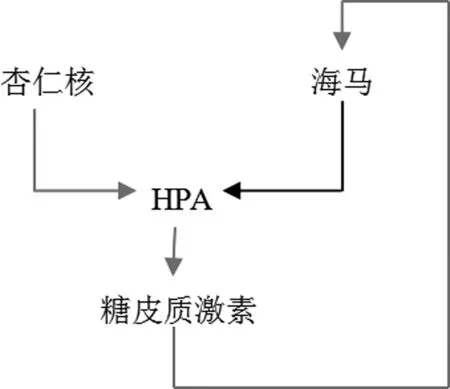
图2 杏仁核与海马区对HPA轴活动的控制(红色表示兴奋性连接,黑色表示抑制性连接)
4 内侧前额叶与HPA轴活动的关系
大脑前额叶皮层与认知功能关系密切,该皮层分为眶部、背部、内侧部和外侧部。内侧前额叶的功能涉及机体控制干扰,抑制与任务无关的表征。其背侧区与各种运动行为相关,其腹侧区与应激情绪及行为反应相关[17,18]。前额叶对杏仁核、HPA轴和脑干蓝斑系统有抑制性下行投射[10]。
在大鼠正常的应激反应当中,内侧前额叶作为皮质醇负反馈作用的靶点,通过与此区域高密度的皮质激素受体相互作用,能有效抑制HPA轴的过度分泌[19]。动物研究表明,毁损大鼠右半球的内侧前额叶腹侧区,可以明显激活HPA轴和自主神经系统的活动,电刺激前额叶可显著改变血浆皮质激素的浓度,说明上述负反馈的作用。
通过功能核磁共振技术,研究者发现抑郁患者的大脑左半球内侧前额叶容量减少,新陈代谢速率降低。这些都说明大脑内侧前额叶皮层在应激反应的调控及抑郁发病可能机制研究方面的重要性。
值得一提的是,Meaney在一项发育学研究当中,对大鼠进行出生后操作:从大鼠一出生起,到出生后21天止,每天将它们与母亲分离15-20分钟。结果发现这些大鼠成年后前额叶的糖皮质激素受体密度显著增加,这意味着前额叶对糖皮质激素的敏感性增加,在额叶水平的皮质激素负反馈变得更加有效,从而抑制皮质激素的过量分泌[18]。
这和我们的经验不符,可以看出,上述实验范式正是经典的早期生活应激模型的建模方法,如果建模成功,一般可以在模型动物身上观察到过高的HPA轴应激反应。这可能与前额叶对应激源的独特反应有关,Diane Diorio等的研究表明,大鼠前额叶对不同种类的应激源(乙醚刺激和束缚应激)反应性有显著不同:同样是内侧前额叶毁损或在前额叶注射皮质激素的大鼠,对于暴露在束缚应激当中的个体,其ACTH和皮质激素的分泌反应增强;而经受乙醚应激的个体则不出现此现象[19],所以单单通过判断前额叶水平的皮质激素负反馈活动是否正常并不可靠,这大概也是在涉及到增强糖皮质激素受体功能的抗抑郁药物时,许多研究只考虑海马区的相关变化,而不参考内侧前额叶区域受体变化的原因之一吧。
5 小结
事实上,涉及到控制及调节HPA轴应激反应的脑区有多个,其中最常被提及的是下丘脑室旁核(HPA轴的直接激活部位)、海马(HPA轴的高位调节中枢)及杏仁核(应激反应执行部位,同时其他脑区对HPA轴应激反应的调节作用亦不容忽视。
许多实验证实HPA轴功能异常与抑郁症有关,因此最近有学者提出,HPA轴可能部分参与抑郁症的发生和发展。所以清楚了解正常状态下HPA轴的活动特点及其调控以及抑郁状态下相关调控发生的变化对探索抑郁症的发病机理意义重大。
猜你喜欢
中国疼痛医学杂志(2022年10期)2022-12-05
磁共振成像(2022年6期)2022-07-30
四川大学学报(自然科学版)(2021年1期)2021-01-26
音乐生活(2020年12期)2020-01-05
电子制作(2019年23期)2019-02-23
中老年健康(2017年9期)2017-12-13
现代计算机(2017年4期)2017-03-29
上海海事大学学报(2016年3期)2016-12-19
校园英语·下旬(2016年4期)2016-05-09
安徽医科大学学报(2015年9期)2015-12-16
