
粉尘螨HSP16-1原核表达体系构建与温度应激响应功能鉴定
2021-04-28 13:36牛栋玲赵亚娥张宛钰郭宏松胡丽
热带病与寄生虫学 2021年2期
牛栋玲,赵亚娥,张宛钰,郭宏松,胡丽
西安交通大学基础医学院病原生物学与免疫学系,陕西 西安 710061
本课题组前期通过转录组测序、qRT-PCR检测和RNA干扰,确认了热休克蛋白家族(HSPs)中有17种基因参与粉尘螨温度应激响应,起关键应激调节作用的基因有6种,即HSF、HSC71、HSP70、HSP83-1、HSP83-2和HSP16-1,且各关键基因调节规律各不相同。HSP70、HSP83-1、HSP83-2和HSP16-1只参与热应激响应,而HSF和HSC71在热应激和冷应激响应中均发挥调节作用[8-10]。该研究第一次在mRNA水平确认了公认的生物应激响应基因HSPs参与粉尘螨抵抗温度胁迫的应激响应。然而,mRNA水平的改变并不能直接反映蛋白质水平的改变。目前虽然能够检索到螨类HSPs原核表达的相关文献[11-13],但都是原核表达体系构建和重组蛋白诱导表达,缺乏蛋白水平的功能验证。
本研究选择粉尘螨HSP16-1为目的基因,尝试建立原核表达体系,探索重组蛋白表达的最佳诱导条件,通过绘制重组菌和空载菌在热胁迫和冷胁迫下的生长曲线,从蛋白水平验证HSP16-1在温度胁迫下的应激响应功能。这些研究结果将为深入探讨粉尘螨HSF、HSP70s和HSP90s等重要HSPs在温度应激响应中的作用机制及生物学功能奠定实验基础。
1 材料与方法
1.1材料
1.1.1螨虫培养与收集实验用粉尘螨来自本实验室人工气候培养箱纯培养,设置温度为(25 ± 1)℃,湿度为70%~75%。振筛分离法获取含螨粉尘[9],采用自制取螨针在显微镜下分离粉尘螨雌性成虫。
1.1.2主要仪器与试剂智能人工气候培养箱和恒温水浴锅(东南仪器,中国);视频显微镜(麦克奥迪,中国);普通PCR仪(Thermo scientific,美国);实时定量PCR仪(Agilent Mx3005P,美国);超声波细胞破碎仪(上海之信,中国);脱色摇床(其林贝尔,中国);扫描仪(Umax,美国);微量RNA 提取试剂盒(Qiagen,美国);逆转录试剂盒、高保真酶PrimeSTAR© HS DNA Polymerase、BamHⅠ、XhoⅠ、T4连接酶(TaKaRa,中国);E.coliDH5α和BL2(DE3)感受态细胞(天根,中国);胶回收试剂盒(OMEGA,美国);BCA蛋白定量试剂盒(鼎国,中国);SDS-PAGE 快速凝胶试剂盒(赫特,中国);预染蛋白Marker(Thermal Fisher,美国);细菌裂解液(行知生物科技,中国);蛋白上样缓冲液(赫特,中国);凝胶成像系统(赛智,中国);电泳仪(伯乐,美国) 。
1.2方法
1.2.1粉尘螨总RNA提取与cDNA合成选取本实验室人工气候培养箱纯培养的粉尘螨,振筛分离法获取含螨粉尘,平铺在干净玻片上,静置几分钟后,于显微镜下挑取活雌性成螨约500只,置于15 mL无菌无酶离心管,液氮研磨破膜。按照微量RNA 提取试剂盒说明书提取粉尘螨雌性成螨总RNA,用反转录试剂盒将RNA反转录为cDNA,-20 ℃保存备用。
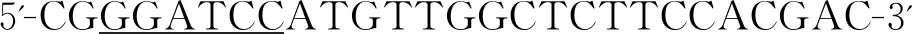
海南琼中抽水蓄能电站上水库副坝一坝址位于一冲沟内,坝基防渗墙厚0.8m,最大墙深约60m,墙底深入至弱风化基岩。副坝一坝址覆盖层分布广,全风化带下限埋深50m~57m,地表有厚近10m的淤泥层,传统防渗墙施工工艺成槽难度极大,在使用液压抓斗配合冲孔桩机施工多次成槽失败后,更换采用德国生产的BC30双轮铣槽机直接铣槽施工,利用其成槽过程中对地层扰动少的优点,顺利实现成槽浇筑,最终按期圆满完成了基础防渗施工。
1.2.3pET32a/HSP16-1重组质粒构建与鉴定 纯化BamHⅠ和XhoⅠ双酶切的HSP16-1 ORF和pET32a载体,采用T4连接酶连接后,转化E.coliDH5α感受态细胞进行克隆。通过Amp(+)LB固体培养基筛选阳性克隆,随机挑取单菌落进行质粒PCR鉴定,阳性克隆送苏州金唯智生物科技有限公司进行测序,通过BLAST比对确认序列及近缘物种。
1.2.4HSP16-1生物信息学分析 用ProtParam软件预测蛋白质组成和理化性质。用ExPASy中的Prot Scale软件进行氨基酸的疏水性分析。蛋白质信号肽用signalP-5.0预测。用NCBI Conserved Domains Search(https://www.ncbi.nlm.nih.gov/Structure/)预测保守区。蛋白亚细胞定位预测使用softberry软件(http://www.softberry.com/berry.phtml?topic=protcom pan &group=programs &subgroup=proloc)。使用在线软件SWISS-MODEL(https://swissmodel.expasy.org/interactive)对其空间结构进行预测。
1.2.5重组蛋白的诱导表达和条件优化 将测序正确的pET32a/HSP16-1重组质粒转化到E.coliBL21(DE3)感受态细胞中,PCR电泳初步鉴定阳性克隆。提取进一步扩大培养,当OD600为0.4~0.6时,用1.0 mmol/L IPTG诱导表达,诱导温度选择16、28和37 ℃[14],诱导时间选择2、4、6和8 h,分别取菌液5 mL。4 ℃10 000×g离心10 min,弃上清,加入1 000 μL PBS重悬洗涤。往重悬菌液中加入细菌裂解液150 μL,冰上静置30 min后,超声破碎细胞膜,超声功率90 W,工作10 s间隙10 s,约5~10个循环,直至液体澄清。参照BCA蛋白定量试剂盒说明书进行蛋白浓度测定,绘制BSA蛋白定量标准曲线,计算样品浓度。加入5×上样缓冲液重悬,金属浴煮沸10 min 使蛋白样品变性,离心收集上清,经10% SDS-PAGE电泳检测分析蛋白表达结果,筛选HSP16-1重组蛋白表达的最佳诱导条件。
1.2.6原核重组表达菌温度胁迫下生长曲线绘制将pET32a和pET32a/HSP16-1菌株各单克隆3个,在液体LB培养基中37 ℃培养,OD600为0.4~0.6时,加入IPTG在最佳条件下诱导蛋白表达,调节各菌液OD值一致,取1%接种到新鲜培养基,37 ℃培养,每隔1 h测定一次各菌液OD值,并记录数据,绘制诱导后37 ℃生长曲线。然后分别在41 ℃水浴锅和-7 ℃冰盐浴处理6、12和24 h,再次1%接种,37 ℃培养,最后绘制热胁迫和冷胁迫下细菌生长曲线。

2 结 果
2.1粉尘螨HSP16-1序列比对与分析 粉尘螨HSP16-1经特异性引物PCR扩增,电泳显示的条带与目的片段大小一致,见图1A;经克隆测序获得462 bp核苷酸序列,比对显示与转录组测序的HSP16-1基因开放阅读框(Genbank:MK896232)完全一致,未发现碱基突变,可编码153个氨基酸,见图1B。核苷酸和氨基酸序列经BLAST比对,覆盖度为100%的分别只比对上一条序列,均为近缘物种屋尘螨α晶体蛋白A链(XM_027339750,XP_027195551),相似度分别为84.63%和87.58%。

注:A为HSP16-1 PCR扩增,M为DL1000 DNA marker,1为HSP16-1 ORF;B为HSP16-1核苷酸氨基酸序列比对,红色字体为预测的保守区;C为 HSP16-1三维结构。
ProtParam分析显示,粉尘螨HSP16-1分子量为17.93 kDa,理论等电点为6.83,带负极残基(Asp+Glu)22个,正极残基(Arg+Lys)22个,总原子数2 506,预测分子式为C786H1249N225O237S9。在编码的153个氨基酸中,含量最多的为Lys(K)9.2%,其次为Asn(N)8.5%、Asp(D)8.5%、Gln(Q)8.5%和Val(V)8.5%。不稳定系数47.98,为不稳定蛋白;亲水性指数GRAVY-0.808,为疏水性蛋白;脂肪指数74.44。蛋白定位综合预测显示HSP16-1定位于细胞核,得分2.5。signalP-5.0预测HSP16-1未发现信号肽。预测的粉尘螨HSP16-1三维结构如图1C所示。
粉尘螨HSP16-1保守区预测,发现氨基酸序列有一个α晶体蛋白HSP23超家族的保守区,共74个氨基酸(在54~127位氨基酸),即NFDVRNFDPNSVQVKLDGNNLQVQAKQEKKADGHYEYREFVRHVTVPQNVQSDQLKCKMDKDGVLRFEAPLKQI;与屋尘螨和梅内欧尘螨相似度高达93%和92%。
2.2重组质粒构建 重组质粒菌液PCR检测显示均有阳性条带,表明转化成功,见图2A。挑取阳性单克隆于37 ℃恒温振荡过夜培养后提取质粒,双酶切成功得到pET32a和HSP16-1两条带,说明原核表达质粒pET32a/HSP16-1构建成功,见图2B。

注:A为粉尘螨pET32a/HSP16-1重组质粒菌液PCR,M为DL1000 DNA ladder,1-12为pET32a/HSP16-1重组菌单克隆1-12;B为双酶切鉴定,M1为1 kb DNA ladder,1为pET32a/HSP16-1重组质粒,2为双酶切pET32a/HSP16-1质粒,M2为DL1000 DNA ladder,白色箭头表示双酶切的HSP16-1 ORF。

注:A为BSA蛋白标准曲线;B为不同诱导条件HSP16-1重组蛋白表达SDS-PAGE分析,M为蛋白Marker,NC为pET32a空载菌。
2.3重组蛋白诱导表达 BSA蛋白标准曲线为y=0.106 3x+0.019 5,R2=0.999 5,说明线性关系良好,能够准确定量待检蛋白浓度,见图3A。HSP16-1重组蛋白可被成功诱导表达,融合蛋白分子量约为35.8 kDa,为HSP16-1(17.9 kDa)与pET32a标签蛋白(17.9 kDa)分子量之和。选择16、28、37 ℃三个温度和2、4、6、8 h四个诱导时间,SDS-PAGE显示37 ℃诱导6 h为最佳诱导条件,诱导效果最好,见图3B。
2.4温度胁迫对HSP16-1重组菌生长的影响 细菌生长曲线显示,pET32a/HSP16-1重组菌和pET32a空载菌在37 ℃未诱导时生长曲线一致(t=1.580,P=0.140 0),见图4A;而在37 ℃诱导6 h后,重组菌随着诱导时间的延长,生长好于空载菌,差异有统计学意义(t=5.334,P<0.05),见图4B。

注:A为37 ℃未诱导时生长曲线;B为37 ℃诱导后生长曲线;无差异表示两组比较,P>0.05;***表示两组比较,P<0.001。
41 ℃分别热胁迫6、12和24 h,pET32a/HSP16-1重组菌和空载菌生长曲线均明显好于37 ℃诱导菌;重组菌生长曲线好于空载菌(t=8.767、6.942、6.320,P均<0.05);细菌生长曲线与胁迫时间有关,热胁迫时间越长,起初1~3 h细菌生长越缓慢,但之后细菌繁殖速度呈快速增长的趋势,见图5。
在-7 ℃分别冷胁迫6、12和24 h,则结果有所不同。冷胁迫后,pET32a/HSP16-1重组菌和空载菌的生长直接受到影响,生长曲线在起初7~8 h内低于37 ℃诱导菌,之后细菌快速生长,高于37 ℃诱导菌;空载菌生长好于重组菌(t=8.874、7.158、5.550,P均<0.05),见图6。

注:A为-7 ℃冷胁迫6 h;B为-7 ℃冷胁迫12 h;C为-7 ℃冷胁迫24 h;****表示两组比较,P<0.0001;***表示两组比较,P<0.001。
3 讨 论
本研究在前期从mRNA水平确认粉尘螨多个HSPs参与温度应激响应的背景下,选择HSP16-1尝试建立原核表达体系,完成了蛋白水平的功能确认。选择HSP16-1作为原核表达的目的基因,其原因主要有:相对于HSP70s、HSP90s和HSF等多个HSP基因,HSP16-1基础表达量高(FPKM为3 864.36),核苷酸序列较短(462 bp),利于PCR扩增测序;相对于HSP16-2在41 ℃热胁迫上调3.27倍,HSP16-1上调4.00倍,调节作用强[10]。经过生物信息学分析,发现粉尘螨HSP16-1属于HSPs中的α晶体蛋白A链亚家族成员,与HSPs的典型结构特征相一致[15]。检索NCBI数据库,目前收录的螨类HSP16s基因序列仅有4条,涉及屋尘螨(Genbank:XP027195551)、梅内欧尘螨(Genbank:OTF73758)、地里纤恙螨(Genbank:RWS26778)和二斑叶螨(Genbank:XP015781289),而粉尘螨HSP16基因未被收录。我们以前期RNA-seq获得的HSP16-1序列为模板,设计特异性引物,经逆转录、PCR扩增和测序,成功获得了粉尘螨HSP16-1全长编码区序列(coding sequence,CDS),全长462 bp。经序列比对,与RNA-seq获得的ORF模板完全一致,没有碱基突变。已投递至GenBank,为今后粉尘螨HSPs基因的研究提供了序列模板。
蛋白质生物学功能的发挥依赖于正确折叠的三维结构,因此对蛋白质三维结构的预测十分必要。常采用计算机模拟来预测三维结构,同源建模是目前比较主流的思路。通过对预模拟的蛋白质序列与PDB数据库中的序列进行相似性搜索,根据相似序列的结构来预测,具有预测速度快、结果准确度高的优点。本研究选择的SWISS-MODEL在线软件基于同源建模方法进行,成功预测出了粉尘螨HSP16-1蛋白质三维结构,为更好地从空间结构的角度研究HSP16-1蛋白的生物学机制奠定了基础。
要实现蛋白水平的功能研究,第一步是要获得重组蛋白。本研究选择大肠杆菌原核表达系统,尝试对粉尘螨HSP16-1进行蛋白表达。大肠杆菌表达系统是最早被采用和目前掌握最为成熟的表达系统,它具有遗传背景清楚、繁殖快、产量高、成本低、IPTG诱导表达相对简便、表达产物容易纯化、稳定性好、抗污染能力强以及适用范围广等优点,是生产重组蛋白最常用的系统[16-17]。载体选择pET32a,是因为pET载体在大肠杆菌原核表达系统中,基础表达水平最低,更利于目标基因的表达。研究设计了3个诱导温度和4个时间梯度,最终筛选出粉尘螨HSP16-1最优诱导表达条件为37 ℃诱导6 h。
近年有多项通过绘制细菌生长曲线以揭示非生物胁迫对玉米、番茄和木薯等植物生长功能影响的研究[18-20]。本研究借鉴这一技术,观察HSP16-1蛋白对重组菌和空载菌在热胁迫和冷胁迫下生长曲线变化,验证蛋白功能。研究发现,热胁迫时重组菌的生长要明显好于空载菌,可能是HSP16-1蛋白在重组菌中被大量诱导表达,发挥热应激响应功能,使得重组菌对热胁迫的抵抗力增强;而冷胁迫时重组菌的生长相对于空载菌较差,说明HSP16-1蛋白在冷胁迫不具有应激响应功能。这与前期HSP16-1 mRNA水平仅在热胁迫上调的结果一致。研究还发现冷胁迫细菌恢复生长要滞后于热胁迫,可能与细菌在低温环境下生理代谢和繁殖速度减慢有关。热胁迫和冷胁迫24 h恢复生长时间要滞后于12 h和6 h,这可能与长时间温度胁迫,导致部分细菌死亡或者活性减弱有关。在温度胁迫后,重组菌和空载菌的生长后期均超过了37 ℃诱导菌,究其原因,可能是除了重组表达的HSP16-1对细菌发挥应激响应作用外,细菌自身的应激响应基因也在发挥作用,使得胁迫后细菌的生长反而好于37 ℃诱导菌。此外,本研究发现重组菌诱导后的细菌生长要低于未诱导,可能是由于IPTG诱导表达的蛋白,对细菌有一定的毒性作用,轻微抑制了细菌的生长[20]。
本研究成功建立了粉尘螨HSP16-1原核表达方法,揭示了粉尘螨HSP16-1在热胁迫具有应激响应功能,建立的方法为在蛋白水平研究粉尘螨等其他螨虫功能基因提供了技术借鉴。
猜你喜欢
中国动物保健(2022年2期)2022-05-05
防爆电机(2021年6期)2022-01-17
江西农业学报(2021年4期)2021-04-20
作文成功之路·小学版(2019年9期)2019-10-17
资源节约与环保(2018年1期)2018-02-08
湖南农业(2017年1期)2017-03-20
中南民族大学学报(自然科学版)(2015年2期)2015-12-16
安徽医科大学学报(2015年9期)2015-12-16
西南医科大学学报(2015年1期)2015-08-22
中国当代医药(2015年9期)2015-03-01
