
广德市笄罩山毛竹根际绿僵菌物种多样性及生态位研究
2021-05-11 10:31陈名君侯囡嵩刘玉军
中国生物防治学报 2021年1期
陈名君,侯囡嵩,林 俨,刘玉军,黄 勃*
(1. 安徽农业大学微生物防治省重点实验室,合肥20036;2. 安徽省科学技术研究院,合肥 230031)
毛竹Phyllostachys edulis是一种地域分布广,经营周期短,经济效益高的优良树种。其具有产量高、用途广和笋材两用等特点,使其具有重要的市场价值,同时毛竹鞭根发达,更新繁茂,防风固沙、保持水土能力强,持续利用时间长,又具有重要的生态价值[1]。毛竹林在中国栽培历史悠久,种植面积广,是我国南方重要的森林类型之一。毛竹在生长过程中受到众多害虫危害,如大竹象Cyrtotrachelus longimanus、竹笋禾夜蛾Oligia vulgaris、竹笋泉蝇Pegomyia phyllostachys、双栉蝠蛾Bipectilus zhejiangensis[2]、筛胸梳爪叩甲Melanotus cribricollis[3]、黄脊竹蝗Ceracris kiangsu[4]等,对毛竹的产量和品质产生严重的影响。
绿僵菌属Metarhizium Sorokin是以金龟子绿僵菌Metarhizium anisopliae为模式种建立的真菌,其有性型在最新的系统分类地位隶属于子囊菌门 Ascomycota、粪菌纲 Sordariomycetes、肉座菌目 Hypocreales、麦角菌科Clavicipitaceae、异虫草属Metacordyceps。2013年Kepler等[5]提出应用MzIGS3基因能够有效将金龟子绿僵菌复合种鉴定到种水平。随后Kepler等[6]将绿僵菌和有性型的虫草联系起来,并分析了绿僵菌属Metarhizium, 普奇尼亚菌属Pochonia,野村菌属Nomuraea和拟青霉属Paecilomyces之间的关系,将异虫草属 Metacordyceps中的很多种重新归入绿僵菌属,包括野村菌属 Nomuraea和最近新描述的属Chamaeleomyces。Luangsa-ard等[7]根据绿僵菌新分类系统,采取多基因分子系统学和经典分类技术结合,在泰国热带森林发现绿僵菌新种M. chaiyaphumense, M. kalasinense, M. prachinense,M. samlanense和M.takense;以及和Metarhizium属相近的新属Nigelia。目前绿僵菌分类系统取得了突破性的进展,这为本研究提供了可靠的技术保证。
有研究表明,绿僵菌对多种毛竹害虫有显著的防治效果。林春穆等[2]筛选出能有效控制浙江双栉蝠蛾的金龟子绿僵菌;杨华等[4]研制出对黄脊竹蝗具有极大防治潜力的绿僵菌微胶囊剂;叶碧欢等[3]测定了平沙绿僵菌对竹林重要地下害虫筛胸梳爪叩甲幼虫具有极显著的趋避作用。近年来越来越多的研究表明,绿僵菌还具有植物内生、植物病害拮抗、植物生长促进和根际定殖等一系列作用。Hu等[8]首次报道金龟子绿僵菌在甘蓝根际具有定殖能力。Wyrebek等[9]研究了加拿大安大略省不同植物根际绿僵菌的分布情况,分离出罗伯茨绿僵菌M. robertsii、棕色绿僵菌M. brunneum和贵州绿僵菌M. Guizhouense 3种绿僵菌,且并非随机分布,而与植物种类具有严格的相关性。
生态位宽度反映了物种利用资源的能力和多样性的大小,空间生态位宽度较高,说明该物种分布区域较为广泛;时间生态位宽度较高,说明该物种适温范围较广[10]。生态位重叠指数反映了不同物种对资源利用程度在空间、时间维度上的相似程度,其值介于0~1,其值越大代表物种生态位重叠越高,当生态位重叠指数大于0.6时,视为显著重叠[11]。研究毛竹根际绿僵菌属真菌物种多样性及在毛竹根际的生态分布,不仅具有重要的理论意义,而且对于保护和促进绿僵菌在毛竹根际的自我维持,更合理的利用该菌控制毛竹林地上地下害虫也具有重要的理论意义。
1 材料与方法
1.1 标本采集
选择素有“竹海”之称的安徽省广德市笄罩山,于2018年秋、冬和2019年春、夏4个季节分别前往采集。在笄罩山海拔100、250和400 m处分别选取阴坡和阳坡设置6个样地,每个样地约100 m2,每个样地内随机选取10株龄期和大小基本一致的毛竹,用铁锹挖出毛竹距表土5~20 cm深处的根系,尽可能地去除根表多余的土壤,然后再装入标记好的自封袋中,带回实验室及时分离处理。在一年的4个季节共采集4次。
1.2 绿僵菌属真菌的分离
毛竹根系用无菌蒸馏水冲洗去除多余的土壤,紧紧附着在根部的土壤可保留。取2 g的根样品,剪成0.5 cm小段,放入装有10 mL蒸馏水的三角瓶中,置于摇床上摇匀1 h。取匀浆样品(100 μL)接种在选择性培养基(20 g/L葡萄糖、200 g/L马铃薯、20 g/L琼脂、0.2 g/L氯霉素、0.2 g/L放线菌酮、0.5 g/L多果腚和0.01 g/L结晶紫)上并涂布均匀,每样品设置3次重复。将已接种培养皿放在25 ℃恒温培养箱中培养,从第10 d开始,陆续从培养皿上挑取出所有真菌菌落,纯化培养。对纯化得到的菌株,通过形态特征鉴定和分子测序,确定其种类[12]。
1.3 形态观察
依据经典分类研究方法,将分离纯化的菌株,接种至1/4 SDAY培养基上,记录菌株直径、菌株颜色质地等参数并拍照。将刚产孢的菌株在光学显微镜下观察产孢结构、孢子形状、菌丝等特征,并记录孢子大小、瓶梗大小和菌丝宽度等,然后进行物种鉴定。
1.4 DNA提取
将纯化并经形态鉴定的菌株,接种到带有玻璃纸的PDA培养基(20 g/L葡萄糖、200 g/L马铃薯、20 g/L琼脂)上,置于25 ℃恒温培养箱中培养3 d后,取0.05~0.1 g菌丝备用。用改良的CTAB法[13]提取基因组DNA。利用核酸测定仪检测基因组DNA质量,并于-20 ℃冰箱保存备用。
1.5 PCR扩增和测序
本试验用于PCR扩增的引物是:MzIGS3-1F(cgt ggc tcc tga cca tgg ttg c)和MzIGS3-4R(gcg ggg gag ccg act tgg a)。PCR 扩增以 25 μL 反应体系进行,其中包括 2.5 μL Buffer,0.5 μL dNTP,正反向引物各 1 μL,0.2 μL Taq 聚合酶,1 μL 模板 DNA 和 18.8 μL PCR 水。测序所使用的引物为:MzIGS3-2F(5′-gtg gct cct gac cat ggt tgc-3′)和 MzIGS3-3R(5′-cgg ggg agc cga ctt gga ttt-3′)。PCR 扩增程序:95 ℃预变性 30 s,95 ℃变性30 s,TM值减去3 ℃~5 ℃退火30 s,72 ℃延伸1 min,循环34次;循环结束后72 ℃延伸10 min。PCR扩增产物在1.5%琼脂糖凝胶上电泳分离,EB染色后于凝胶成像系统检测拍照并送至北京六合华大基因科技股份有限公司测序。
1.6 生态位宽度和重叠

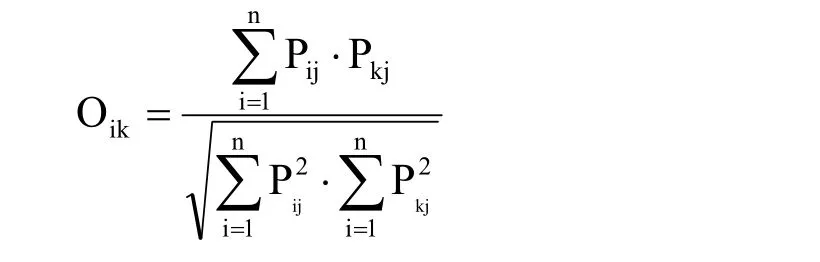
式中,Pij=nij/Nij即种i在第j个资源状态下的个体数占该种所有个体数的比例。其中,在计算时间生态位宽度、时间生态位重叠时,j表示某个调查月份;在计算空间生态位宽度、空间生态位重叠时,j表示某个样地。R为总样地数或总调查月份次数。
2 结果与分析
2.1 绿僵菌属真菌分离鉴定结果
通过对安徽省广德市笄罩山毛竹根系绿僵菌的采集分离,经形态特征初步鉴定,共从毛竹根际分离出164株可能为绿僵菌属真菌的菌株。经提取DNA并进行PCR扩增后,将测序得到的序列在GenBank数据库中进行同源性比较。经鉴定共获得绿僵菌163株,其中罗伯茨绿僵菌的多度最高,为98株,相对多度为0.601;其次为一种待鉴定绿僵菌,多度和相对多度分别为47和0.288,平沙绿僵菌有11株,贵州绿僵菌最少,仅有7株。4种绿僵菌菌落形态、产孢结构和孢子形态详见图1。
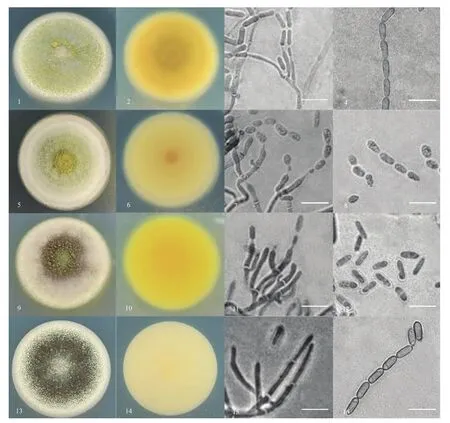
图1 四种绿僵菌在SDAY/4培养基上菌落形态、产孢结构和孢子形态(标尺=10 μm)Fig. 1 Colony morphology of four Metarhizium species on SDAY/4 and mature conidiogenous cells and conidia (Bar=10 μm)
2.2 毛竹根际绿僵菌属真菌空间分布情况
图2结果表明,从绿僵菌的海拔分布来看,海拔100和400 m处的毛竹根际绿僵菌属真菌多样性明显高于海拔250 m处。在设置的3个海拔梯度中,海拔100 m处分离得到的根际绿僵菌菌株最多,其中罗伯茨绿僵菌为优势种;其次是400 m处,优势种是一种待鉴定绿僵菌;250 m海拔处分离得到的根际绿僵菌菌株最少。罗伯茨绿僵菌在毛竹根际的数量随着海拔的升高而递减。一种待鉴定绿僵菌和平沙绿僵菌在毛竹根际的数量在海拔250 m处最低,而贵州绿僵菌的数量则随着海拔的增高而略有上升。
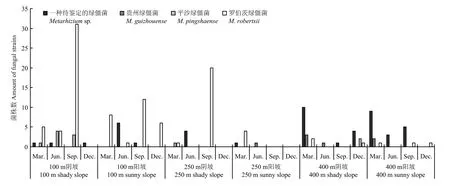
图2 毛竹根际绿僵菌空间分布情况Fig. 2 Summary of the spatial distribution of the Metarhizium species in the rhizosphere of Phyllostachys edulis
从绿僵菌的坡向分布来年,阴坡处的毛竹根际绿僵菌多样性高于阳坡处。4种绿僵菌中,平沙绿僵菌只在阴坡处分离得到,可能有较强的坡向偏好。其他3种绿僵菌均未发现有较明显的坡向偏好。
2.3 毛竹根际绿僵菌的时间分布情况
研究一年中4个季节毛竹林根际绿僵菌分布情况,发现9月绿僵菌最多,其中罗伯茨绿僵菌为优势种;3月分离得到的绿僵菌中一种待鉴定绿僵菌和罗伯茨绿僵菌较多;6月分离得到的绿僵菌中最多的是一种待鉴定绿僵菌;12月分离得到的绿僵菌最少。由图3可以看出,除罗伯茨绿僵菌外,其余3种绿僵菌在毛竹根际的数量随季节的变化而递减,而罗伯茨绿僵菌在夏季毛竹根际分布达到峰值。
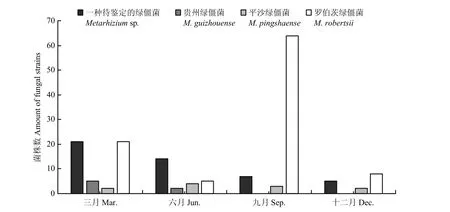
图3 毛竹根际绿僵菌时间分布情况汇总Fig. 3 Summary of the temporal distribution of Metarhizium species in the rhizosphere of Phyllostachys edulis
2.4 毛竹根际绿僵菌时空生态位宽度
同种绿僵菌的空间生态位宽度呈现明显的季节变化。一种待鉴定绿僵菌和贵州绿僵菌在春、夏季节中,空间生态位宽度较高,在秋季和冬季的各月中则较低;罗伯茨绿僵菌在春、秋季中,空间生态位宽度较高,在夏、冬季中则较低;而平沙绿僵菌仅在春季生态位宽度略高。罗伯茨绿僵菌的空间生态位宽度在春、秋、冬三季中均为最高的,然而在春季一种待鉴定绿僵菌的空间生态位宽度最高。总体来看,春季中毛竹根际的绿僵菌空间生态位宽度最高,4种绿僵菌中,贵州绿僵菌的时间生态位宽度最低,其余3种绿僵菌的时间生态位宽度差异较小,其中时间生态位宽度最高的是一种待鉴定绿僵菌(表1)。
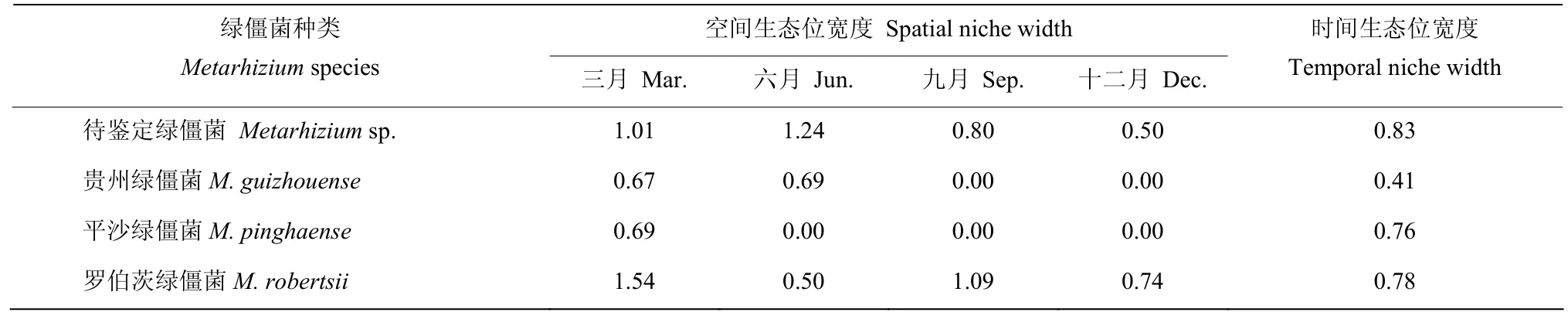
表1 毛竹根际绿僵菌的时空生态位宽度值Table 1 The spatial niche width and the temporal niche width of Metarhizium species in the rhizosphere of Phyllostachys edulis
2.5 毛竹根际绿僵菌时空生态位重叠
不同绿僵菌间的空间生态位重叠值呈现一定的季节变化。在春季,一种待鉴定绿僵菌和贵州绿僵菌的空间生态位重叠值最高;而在夏季和秋季中,平沙绿僵菌和罗伯茨绿僵菌的空间生态位重叠值最高;在冬季中,待鉴定绿僵菌和平沙绿僵菌空间生态位重叠值最高(表2)。
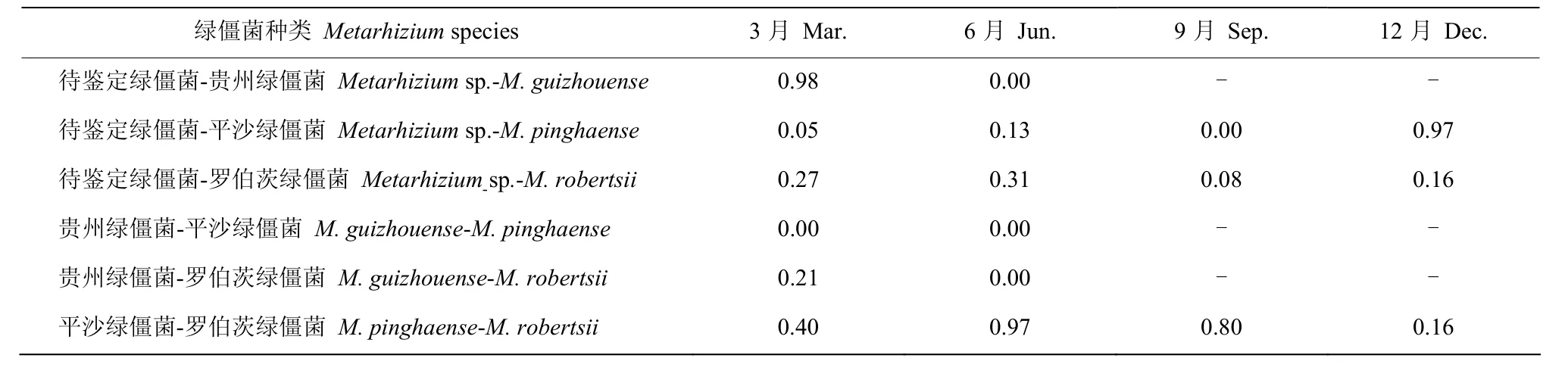
表2 毛竹根际绿僵菌的空间生态位重叠值Table 2 The spatial niche overlap of Metarhizium species in the rhizosphere of Phyllostachys edulis
不同绿僵菌间的时间生态位重叠值差异较大,分布在0.31~0.92。其中待鉴定绿僵菌和贵州绿僵菌的时间生态位重叠值最高,而贵州绿僵菌和罗伯茨绿僵菌的时间生态位重叠值最低(表3)。

表3 毛竹根际绿僵菌的时间生态位重叠值Table 3 The temporal niche overlap of Metarhizium species
3 讨论
本文从毛竹根际共分离出4种绿僵菌,分别是罗伯茨绿僵菌、平沙绿僵菌、贵州绿僵菌和一种待鉴定绿僵菌,其中罗伯茨绿僵菌的数量最多,其次是待鉴定绿僵菌。根据现有报道发现,美国和加拿大所分离出的根际绿僵菌中,罗伯茨绿僵菌均是分布最广泛的,与本文的研究结果一致[16,17]。但在研究日本筑波县5月和6月森林不同植物根际绿僵菌时,发现优势种为平沙绿僵菌[18],其差异可能是由于研究季节较单一,未能全面反映一年中植物根际绿僵菌分布情况。另外,也可能与植物种类、土壤环境、植被坡向等因素有关。
周芳纯等[19]研究发现,夏末、秋初和初春是毛竹的快速生长时期。在本实验中3月和9月分离出大量的罗伯茨绿僵菌,明显多于另外两个月,然而其他几种绿僵菌则没有表现出此种情况,说明罗伯茨绿僵菌可能与毛竹生长有一定的协同或者促进关系。Sasan和Bdochka[20]研究发现罗伯茨绿僵菌的孢子能粘附在柳枝稷和菜豆根表,并能促进其根系生长。Liao等[21]研究发现罗伯茨绿僵菌作为根际微生物促进了植物对氮的吸收并增强了植物抗病性,同时罗伯茨绿僵菌寄生昆虫后,可将从昆虫中吸收的氮营养物质传给植物,还可分泌各种有机酸溶解土壤中的固态磷和二氧化碳[22]供植物生长发育之用,且可以消除土壤中的铅污染,也能帮助植物吸收土壤中的铁和锌[23]。综上所述,可以发现罗伯茨绿僵菌确实对植物生长有促进作用。
从毛竹根际绿僵菌的时空分布情况可以看出,根际绿僵菌的优势种在不同海拔和不同季节不完全相同,因此无法证明绿僵菌和植物之间存在特定的相关性,这一观点与日本学者Nishi等[18]研究结果一致。根际优势绿僵菌可能是由植物生长环境因素综合决定的。Bidochka等[24]研究发现绿僵菌对植物种类的偏好可能是由于其对热生长偏好的差异以及对紫外线辐射的适应能力而不是对植物物种本身的适应性。因为他们所分离出的罗伯茨绿僵菌与棕色绿僵菌相比更喜欢高温,并且对紫外线辐射具有更高的抵抗力。Fisher等[25]同样认为生境才是决定绿僵菌种类的原因,而不是植物种类。他们研究中所采集的圣诞树根际绿僵菌种类与其他三种植物根际的绿僵菌种类明显不同,因为他们采样点相比其他植物的采样点的区域范围更小。本文仅研究了毛竹根际绿僵菌分布情况,且在分离过程中可能存在一定的误差,要想进一步证明绿僵菌和植物是否存在特定的相关性,今后还需要进一步对不同植物的根际绿僵菌进行更为详尽的研究。
猜你喜欢
湖北植保(2022年5期)2022-12-24
意林·少年版(2020年13期)2020-08-02
中国新闻周刊(2018年22期)2018-06-20
西部林业科学(2018年2期)2018-05-16
茶叶学报(2018年4期)2018-02-28
———“人民的选择”
海外星云(2017年9期)2017-05-22
福建农业科技(2015年1期)2015-02-27
福建农业科技(2015年1期)2015-02-27
电影故事(2015年26期)2015-02-27
生物技术通报(2014年2期)2014-04-08
