
以晶体生长原理重塑蚕丝柔性材料世界的未来
2021-05-20 10:08刘向阳
人工晶体学报 2021年4期
邱 武,刘向阳
(1.厦门大学海洋与地球学院,近海海洋环境科学国家重点实验室,厦门 361005;2.新加坡国立大学物理系,新加坡 117551)
0 晶体、非晶体与柔性材料
自然界的物质,常以四种形态存在,即晶态、液态(包括其他非晶态)、气态与等离子态。其中,晶态与非晶态属凝聚态。晶态,指内部粒子(原子、离子或分子)在空间按照一定规则周期性重复排列构成(或长程有序)的物质;而非晶态,指其内部的原子或分子的排列则呈现杂乱无章的分布状态,或只有近程有序的物质。常见的非晶体包括液体、无定形体和玻璃体等。在适宜的条件(如生长速率合适时),晶体能够自发地呈现封闭的规则和典型的凸面多面体外形。在晶体内部由于各个方向排列的内部粒子之间的距离不同,导致晶体往往在各个方向表现出的物理性质(如硬度、导热性和光学性质等)存在差异,即所谓的各向异性。对于典型的晶体,具有固定的熔点,而对于非晶体,则没有固定的熔点。晶体在现代社会,起着举足轻重的作用,如激光、非线性晶体,硅半导体,金刚石,石墨(烯)等。
近些年来,柔性材料,包括各种聚合物、水凝胶、天然与合成纤维、蚕丝等,从更广泛的意义上来说,在人们生活,以及生物医药方面,起着不可替代的作用,并越来越引起人们的重视[1-9]。特别是近年来发展起来的柔性电子,将影响着大健康、人工智能等方面的发展[10-12],而柔性电子的基础,在于柔性材料。

图1 晶体、非晶体与柔性材料的关系,柔性材料是晶体和非晶体两种形态的混合体Fig.1 Correlation between crystals, amorphous and soft materials, soft materials are combinations of crystals and amorphous
柔性材料原先被认为是非晶材料。然而,越来越多的实验现象证实,包括蜘蛛丝、蚕丝、蛋白质凝胶等在内的许多柔性材料,内部存在着典型的晶体和非晶体(见图1)。这些柔性材料,既可以对X-射线衍射实验做出反应,意味着内部存在晶体;同时又在介观尺度上呈现出与典型晶体显著不同的性质,意味着内部并不完全由晶体充斥。最新的研究表明,对于很多柔性材料,它们内部的基本结构单元是晶体网络[13-19]。在这样的晶体网络中,微晶通过不同的方式连接组成晶体网络结构。这种连接方式,既可以是微晶本身,也可以是其他晶体,还可以是非晶态的分子甚至无定形的分子链。介观晶体网络结构,对柔性材料的宏观性能起着举足轻重的作用。美国能源部《从量子化到连续介质:介观科学的机遇》的白皮书中指出:“软物质宏观的性能,明显来源于介观尺度结构,而非原子或纳观尺度结构”,明确地界定出介观结构在影响柔性材料宏观性能方面的作用[13,15,17]。
基于以上事实,了解柔性材料的介观结构特别是晶体网络结构与宏观性能的关联,就显得越来越重要。在柔性材料领域,大量实验结果证实,在材料内部,晶体网络的拓扑结构、相关长度、对称/有序性度、关联强度等四个因素,直接决定了介观材料的性能(见图2)[13-16,20]。迄今为止,晶体网络的概念已得到同行的广泛认可,现在普遍认为“由组装引起的晶体生长形成的晶体网络,是一种实现生物材料结构的重要途径”[15,18]。

图2 (a)柔性材料介观多级结构模型[13];(b)柔性材料介观结构的宏观性能表现,呈现特定的反映,具体来说,这种表现通过四个结构要素来表现:晶体网络的拓扑结构、相关长度、对称/有序性度、关联强度[13-16,20]Fig.2 (a) Mesoscopic hierarchical structures of soft materials[13]; (b) the macroscopic performances of mesoscopic structure soft materials are highly correlated to four structural factors, they are: the topology of crystal network, the correlation length, the ordering and the interaction[13-16,20]
对软物质的研究,始于一百多年前。学术界对相关课题已经进行了广泛研究。一方面,更新了许多新的软物质系统的相关知识;另一方面,基于相关物理学、生物学和化学领域的新需求,还发展出了许多关于软物质的新应用。软物质系统具有显著的热波动、介观多尺度自组装结构、熵驱动有序无序转换、宏观柔性等共同特征。简言之,这些系统具有“小刺激、大反应”和强烈的非线性。
在不同类型的软物质系统中,柔性材料由于在大型健康及相关领域的广泛应用,近年来被列为最重要的材料之一。结合大数据和人工智能技术,柔性材料和相关的柔性电子器件将重塑人类的生活方式和工作风格。目前社会的主要关注点是识别和制造高性能的新柔性材料,用于制备柔性光子/电子设备和环保智能的组织材料,应用于化疗催化、药物输送、基因传递、生物成像和组织工程、远程诊断等相关领域。特别是可穿戴、可植入、可生物降解/可吸收和可注射的柔性器件将对人类健康和日常生活产生巨大影响。
在远程医疗与大健康方面,柔性电子器件与柔性传感器起着关键性的作用。柔性电子器件的性能,在很大程度上取决于构成柔性电子器件的柔性材料的各项特征。此外,作为柔性传感器,其中的柔性材料应对生物友好或对人的皮肤友好。因此,确定满足制造具有理想性能和效益的柔性电子器件所有要求的材料是一项重大挑战。显然,这是寻找合适柔性材料应用于柔性电子研究的主要障碍之一。目前从柔性材料到柔性电子器件的最新策略,是首先将柔性材料进行介观结构重构以实现介观功能化,再将其制成柔性介观电子/传感器件(见图3)[9]。

图3 基于丝蛋白的柔性材料发展之路。在丝蛋白柔性材料的形成过程中通过控制结晶等结构形成动力学,可以实现对其介观结构的重构,并通过介观掺杂实现介观功能化,制备丝蛋白柔性介观功能材料。对这样的介观功能材料进行后续的介观刻蚀等介观制备工艺,便可以进一步制备基于丝蛋白的介观柔性光电器件[9]Fig.3 Developing road of silk fibroin-based soft materials. By controlling nucleation pathways and corresponding kinetics, the mesoscopic structures of silk flexible materials can be reconstructed. Afterwards, via mesoscopic functionalization, silk mesoscopic functional materials are created. Via meso lithography and other fabrication technologies, such flexible functional materials can beappplied for synthesis of next generation of silk mesoscopic electronics/photonics devices[9]
在本文中,将主要研究在丝蛋白(SF)发生折叠/重新折叠的过程中通过改变路径,实现丝蛋白材料的介观结构调控,并赋予其不同而广泛的全新功能。在这方面,还概述了一项创新材料战略,即制造非同寻常的多功能柔性材料,应用于建造性能优异、应用广泛的柔性电子器件。
作为一种日益重要的柔性材料,蜘蛛丝、蚕丝纤维及再生丝蛋白材料由于其优异的生物相容性与可控生物降解速度,而受到日益广泛的关注。蚕丝纤维材料的产量丰富,而且还具有优异的机械性能、光学性能和电学性能,这使其成为下一代生物相容柔性电子器件的重要研究热点[21-23]。由于其良好的体外和体内生物相容性、优秀的力学性能和可控的生物降解速度,蚕丝纤维在生物医学、组织工程应用领域,已经受到广泛的重视[3-6,13,15]。丝蛋白制成的生物医制品,不仅可避免人类染上克雅二氏(人类形式的疯牛病)的风险,同时也是美国食品和药物管理局(FDA)批准的医用材料。将蚕丝蛋白溶解、提纯、功能化之后再制备成薄膜、凝胶、小球、纤维、多孔支架等各种形式的再生蚕丝蛋白超分子材料,可以极大地扩展蚕丝的应用范围。
另一方面,丝蛋白材料在优秀的具有生物相容性的柔性光电子器件方面表现出了很大潜力,受到越来越广泛的关注。迄今为止,经介观掺杂实现其功能化调控的丝蛋白材料,已经被应用于多种柔性光电子器件,如,作为OFET(organic field effect transistors)中的介电层,利用忆阻特性制备存储器等[24]。因此,基于丝蛋白材料的功能化,能发展出一系列基于丝蛋白材料的具有生物相容性好、降解可调的光电元器件。而在可穿戴、可植入、可生物降解/吸收、可注射的柔性器件方面,丝蛋白材料同样将对人类未来的健康、生活产生巨大影响[9-12]。
蚕丝和丝蛋白材料具有典型的多级晶体网络结构,主要涉及了二级结构(β折叠),微晶和微晶网络[13,25]。这里,值得思考的问题是,在柔性材料中,为什么存在晶体网络?这个问题应该从以下两个方面进行理解:(1)晶体网络在形成“巨大”的、相互连接的三维空间结构过程中,其“编织”网络或网络扩张的方式,是比较迅速与随意的,它们不仅可以如晶体一样,从空间某些点生长出来;也可以由几个局部小网络通过相互渗透连接而成[13,25]。(2)柔性材料的许多物理性质,是由分子间非化学键(如氢键、疏水相互作用、范德华力等多种形式)的弱相互作用决定。而以晶体有序的形式,组合成的弱相互作用“集”,具有与化学链相比拟的强度与稳定性。举例来说,尽管单个氢键的断裂能相比于化学键要弱得多,但丝蛋白材料内相对的两条分子链间往往由于存在数十个氢键,在这些氢键的协同作用下,分子链间的相互作用“集”便可以得到显著增强(见图4)[15]。事实上,正是由于各种弱相互作用的协同合作,柔性材料的稳定性才得以保存[17]。这里,蜘蛛丝牵引丝线所有的超高强度,就是一典型的例子[26]。在蜘蛛丝牵引丝内部的结晶网络内,由微晶作为网络的结点,在晶体网络受到外力拉伸时承担了大部分应力。破坏微晶,需要克服微晶内存在的β折叠间的疏水相互作用“集”,而破坏每一个β折叠,更进一步需要克服β折叠内β链间的氢键相互作用“集”[15,17]。

图4 丝蛋白材料中微晶内各种相互作用的强度[15] (a)不同类别相互作用的强度估计;(b)β结晶导致的强度增强Fig.4 Summary of variety of interactions involved in SF materials and enforcement strength in crystallization (crystallization force)[15] (a) table of interaction strength of different categories of interactions; (b) illustration of thestrength enforcement due to β-crystallization
介观晶体网络的基础是晶体,由于晶体网络的重要性,对晶体生长进行研究的方法,包括研究其结晶动力学等,也可应用到研究具有典型晶体网络结构的柔性材料之中。具体来说,本文主要关注柔性材料中晶体网络形成的相关动力学,通过介观调控实现结构的介观重构,同时通过引入不同的丝蛋白分子成核引发剂(种子)或通过介观掺杂的方法实现丝蛋白材料的介观功能化。最后再对丝蛋白柔性功能材料实施包括介观刻蚀在内的介观制备工艺,便可以发展出全新的基于丝蛋白材料的柔性介观光、电器件[10]。因此,晶体生长或结晶动力学的知识重要性,不只局限在传统的晶体生长领域,它还在柔性材料的研究领域,起到意想不到的关键性作用[11]。
在本综述中,首先详细讨论了丝蛋白材料的介观多级结构和晶体网络结构,且着重说明了各级结构特别是晶体网络结构中相关的结构单元对其宏观性能的影响。由于晶体网络的重要性,本文还简要介绍了对丝蛋白材料进行介观结构表征的具体技术手段。由于丝蛋白材料的制备符合蛋白质分子的结晶原理,接下来,从成核和结晶的角度,详细讨论了涉及调控丝蛋白分子折叠的路径,丝蛋白材料介观结构的重构方法及控制机制。最后,为了验证晶体生长理论的正确性,还以丝蛋白薄膜为例,说明了晶体生长理论可以应用于制备其他种类的功能化丝蛋白材料。
1 丝蛋白材料的介观多级结构和晶体网络
1.1 丝蛋白材料的介观多级结构
由于丝蛋白材料的介观结构对其宏观性能起着直接甚至决定性的影响,对丝蛋白材料的介观结构,特别是晶体网络结构进行深入且准确的表征就显得尤为重要。与此同时,掌握丝蛋白材料中晶体和晶体网络结构形成的机理,从而应用于精确控制与介观构筑丝蛋白材料的晶体网络结构,就可以构建功能全新的丝蛋白材料,并将之推广到其他种类的柔性材料中。如通过介观重构,可将原先不发光的柔性材料转化成发光柔性材料,也可将原先不导电的柔性材料转化成导电的柔性材料[11]。
虽然对蜘蛛丝和蚕丝的应用和研究已有相当长的时间,但由于表征手段的限制,迄今为止对其内部结构存在许多混淆的认识。随着介观结构和晶体网络结构的概念深入人心,在大量实验基础上,不同形式的丝蛋白材料都被证实,其内部存在典型的纳米微纤,而随着技术发展,原子力显微镜谱第一次被用来证实纳米微纤实质是典型的纳米微晶采取“渔网”结构而构成的晶体网络[27]。结合相关实验结果和模拟结果,本课题组第一次提出了以下丝蛋白材料的五级介观多级结构[13]。自下而上,这五级结构分别为:氨基酸序列、二级结构(β折叠)、微晶、微晶网络(纳米微纤)和纳米微纤网络(见图5)。其中,β折叠是后续微晶和微晶网络的基础,它的含量主要由蛋白质的氨基酸序列决定。一般来说,微晶由于其力学性质相对稳定,在丝蛋白材料受到外力拉伸时,可以承担晶体网络内最多的应力,在微晶没有被破坏前,丝蛋白材料可以有效地保持结构完整。实际上,当丝蛋白材料内部没有形成微晶时,这样的丝蛋白材料不仅会轻易地溶解于溶液中,还往往表现出令人失望的力学强度。除了微晶,晶体网络中微晶密度和微晶取向度,则是另外两个与丝蛋白材料力学性能直接相关的重要结构参数[15]。
1.2 丝蛋白材料内的晶体网络
显然,丝蛋白材料的晶体网络结构主要涉及的是三级结构(即β微晶)和四级结构(即β微晶网络),考虑到β微晶由众多的β折叠通过分子间相互作用力互相堆砌而来,二级结构也被考虑为丝蛋白材料晶体网络的组成部分。值得注意的是,并非所有的β折叠都可以形成微晶。实际上,对丝蛋白材料中的β构象总量和结晶度(即微晶含量)进行量化后证实,β构象的总量总是高于结晶度,因此可以得出以下结论:β构象总量不仅包括形成微晶的分子间β折叠,还包括很多普通的不能形成微晶的分子内β折叠。之所以认为β折叠含量和β微晶的含量都直接与丝蛋白材料的宏观力学性能相关,是因为已经证实,不同丝蛋白材料内的β折叠含量和结晶度总是存在着显著差异,而正是由于这些差异,导致了不同丝蛋白材料迥异的力学性能。举例来说,在蜘蛛的牵引丝纤维中,β构象总量和分子内β折叠含量分别为51%和29%。而在柞蚕丝纤维中,分子内β折叠含量仅为23%。至于桑蚕丝的分子内β折叠含量,相比之下它的比例要小得多,仅占9%。相应地,反映到力学强度上,蜘蛛牵引丝的力学性能可以高达1.3 GPa,远远高于蚕丝的300~400 MPa[15]。
利用X射线衍射技术对天然桑蚕丝纤维的微晶结构进行表征时发现,该微晶具有典型的正交晶格(空间群为P22121)结构,单元尺寸分别为a=0.92 nm、b=0.94 nm和c=0.694 nm,其中a是微晶内沿着β折叠堆叠的方向;b是微晶内β折叠中氢键的方向;c是微晶中β链的方向[15]。最近,经实验证实,微晶的尺寸可以影响微晶的稳定性,继而影响丝蛋白材料的宏观性能。具体来说,蜘蛛牵引丝纤维中的微晶尺寸,特别是沿c方向(即纤维长轴方向)的尺寸,在一定程度内(如,大于6 nm)越大时,该微晶会表现出更强的稳定性。至于微晶在b方向上的尺寸,它们对微晶稳定性的影响恰好相反,即,当微晶在b方向上的尺寸较小时,往往这样的微晶具有更高的强度、刚度和韧性。实际上,当微晶的b方向尺寸限制为2~4 nm时,可以取得最佳的稳定性。从结晶学的角度来看,这种尺寸效应,可以如下解释:与小微晶相比,大微晶内往往还会有更多的缺陷或位错,这会降低大微晶的力学稳定性[13]。由于大晶体的形成通常依赖于具有一定有序度或对称性的组装,理想情况中,在晶体的生长过程中,从溶液相中的结晶单元通过外延生长转移到晶核上,只有经历高度有序的组装才能生长为结构完美的晶体。然而,在某些特定条件(如,溶液的过饱和度太高以至于发生失配的外延生长的势垒过低)下,晶核和后续生长单元的界面上会发生位错。这时,新形成的生长层将会偏离原本晶核的结构,导致最终生成的晶体内具有显著的位错或缺陷,因此导致其稳定性反而降低[15]。
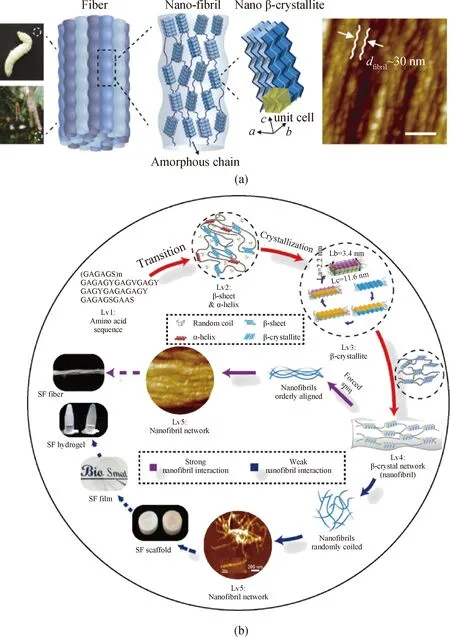
图5 (a)蜘蛛丝蛋白/蚕丝蛋白内部存在典型的晶体网络结构[14];(b)丝蛋白材料的介观多级结构[15]Fig.5 (a) Schema of the crystal network structures within soft materials including silkworm silk fiber and spider fiber[14]; (b) mesoscopic hierarchical structures of SF materials[15]
尽管丝蛋白通过结晶变成微晶对提升丝蛋白材料的力学性能很重要,但形成单独的微晶,仍不足以赋予丝蛋白材料超强的力学性能。事实上,微晶作为一种结构稳定的单元,将众多的微晶联系起来构成网络结构的方式,能够更大程度上决定丝蛋白材料的性能。最近,原子力显微镜已证实,丝蛋白材料内部的微晶网络,采取了所谓的“渔网”状的拓扑结构[27]。即,由无定形链将众多微晶联系起来而形成的网络。该网络中,由于微晶性质稳定,在“渔网”中可以被视作结点,在纳米微纤被拉伸过程中承担主要的拉力。而无定形链的存在,又赋予了纳米微纤一定程度上的柔性。通过采用这种拓扑网络结构,纳米微纤获得了自然界中相对最柔韧、最坚固的力学性能。通过蒙特卡洛方法模拟当丝纤维采取不同晶体网络的拓扑结构后发现,选择β微晶作为纳米“渔网”的交联点的结晶网络结构时,相关的丝纤维的强度能比淀粉状蛋白样结构和独立未交联的分子链结构(“板段”结构)高1 000倍[27]。
由于晶体网络中众多的微晶通过“渔网”状的拓扑结构联系在一起,当这样的网络在整体受到外力拉伸破坏时,应力可以绕过被破坏的微晶从而转移到周围尚未断裂的微晶上,促使更多的相邻微晶参与到承担外力的任务中。因此,当微晶网络内的微晶数目更多(换言之,微晶密度更高时),这样的微晶网络理所应当具备更强的力学稳定性。比较蜘蛛牵引丝和蚕丝纤维的结构时发现,虽然蜘蛛牵引丝的结晶度仅为蚕丝纤维的一半,但由于蜘蛛牵引丝中微晶的尺寸仅为蚕丝纤维中微晶的25%,通过简单的计算可以得知,蜘蛛牵引丝内的微晶密度约为蚕丝纤维中的两倍。显然,这一微晶密度的差异,可以解释为何蜘蛛牵引丝的力学强度远高于蚕丝纤维[13]。除了微晶密度,由于晶体网络中微晶的长轴方向并不总与纤维轴向保持一致,所以微晶的取向也在很大程度上影响最终晶体网络的性能。具体来说,当微晶和纤维轴向保持比较一致时,撕裂微晶需要克服的氢键较多;当取向不一致时,仅需要破坏少量的氢键便可以使微晶的稳定性被破坏。换言之,取向较好的微晶网络更加稳定[15,27]。
通过上述针对丝蛋白材料介观多级结构的介绍,可以得知晶体网络的重要性,尤其涉及微晶的形成,包括如何控制后续微晶网络中的微晶密度和取向度,对于设法提高丝蛋白材料的宏观力学性能起着至关重要的作用[13]。鉴于丝蛋白材料中涉及结晶网络的结构参数(即,二级结构中β折叠的含量,三级结构中β微晶的含量及四级结构中β微晶的密度和取向度)的重要性,对它们进行准确的量化表征就显得至关重要。在本综述中,将简要介绍几种针对柔性材料中与晶体网络相关的常见结构表征技术。
2 表征丝蛋白材料中晶体结构和晶体网络结构的技术
对于柔性材料,特别是带有晶体网络的丝蛋白材料中的结构进行表征,其常规方法主要包括傅里叶变换红外光谱(FTIR光谱)、拉曼光谱、圆二色谱(CD)和X射线衍射谱。
由于其简单和实用的特点,FTIR光谱技术成为了研究包括丝蛋白材料及其他蛋白质基材料内部二级结构含量最早也最广泛应用的方法。FTIR信号经过解卷积操作后,光谱可以被分解成包含有β折叠、α螺旋、β转角和无规则卷曲四种二级结构的多个高斯峰拟合。其中,β折叠的特征峰位于1 620 cm-1和1 698 cm-1附近,而非晶成分的特征峰则位于1 645 cm-1附近。每种二级结构成分的含量可以通过测量对应特征峰的面积比值来定量确定。除FTIR光谱外,拉曼光谱是另一种可无损表征丝蛋白材料二级结构的技术。一般而言,拉曼光谱可以提供与FTIR光谱分析互补的信息。与FTIR和拉曼光谱技术相比,圆二色谱主要用于测定丝蛋白分子链在溶液中的构象(蛋白质二级含量)及其变化过程。由于不同的二级结构所产生的CD谱带位置和强弱都不相同,因此可以通过CD光谱来研究丝蛋白在溶液中的构象。
X射线衍射(XRD)技术则是直接对晶体网络进行定量测量的最常用的技术。通过测量X射线碰撞位于晶体网络中的微晶发生衍射后在空间的分布以及测量信号所在的方位和强度,便可以反推出内部的晶体网络的相关结构。其中,广角X射线衍射(WAXD)/广角X射线散射(WAXS)技术被广泛用于表征晶体网络中微晶的尺寸大小和取向度,及定量测量微晶含量(结晶度)。而小角X射线散射SAXS技术则被用于测量蚕丝和蜘蛛牵引丝内晶体网络中相邻微晶间的距离[15]。
3 丝蛋白分子折叠与丝蛋白材料的介观重构及控制机制
3.1 丝蛋白材料的“自组装机理”:“液晶”和“胶束”模型
由可溶性蛛丝蛋白或丝素蛋白形成固体的丝纤维(及其他非纤维状的柔性材料)的过程,是一个复杂的生化和物理变化过程。在蜘蛛体内,这个过程发生在特殊的器官,即“纺体”中(桑蚕体内对应的器官则为丝腺)。迄今为止,已有多种模型被提出用于阐释蜘蛛牵引丝在“纺体”内的产生过程。其中最有名的两种模型分别是“液晶”模型和“胶束”模型。为了详细阐述这两种模型,首先简介下“纺体”的结构。蜘蛛“纺体”主要由三部分组成,即腺体、导管和纺丝头。以蜘蛛的大囊状腺体为例,它的腺体是一个单独的大囊,大囊尾部有一段长尾,在长尾内会分泌蛛丝蛋白。大囊的另一端,连接了一根直径由粗变细的导管,导管分成了三个“之”字形分支,丝蛋白在导管中的浓度,从第一分支向第三分支递增,最终达到20%~30%(质量分数)的浓度。在导管的末端,连接着所谓的纺丝头,纺丝头内存在一个“阀门”,它像一个夹子一样抓住刚刚开始固化的初生纤维,通过挤出初生纤维内多余的水分,最终使完全固化的丝纤维在纺丝头中被拉出[15]。
早先,利用偏光显微镜对蜘蛛的导管和腺体进行研究后发现,储存在腺体内和导管前部的蛛丝蛋白溶液,表现出了典型的液晶结构,这也是“液晶”模型名称的由来[27]。该模型认为,当蛛丝蛋白刚刚在腺体后部被合成并分泌时,此时的蛋白分子链仍处于紧密缠结的状态,形象地说,处于螺旋结构的分子链段紧密排列成棒状。这些蛋白分子链,逐渐组装成向列相的棒状聚集体,且这样的棒状聚集体仍保持与腺体的分泌壁垂直的方向;当蛛丝蛋白离开腺体进入导管后,分子链的方向开始变得与导管流向平行。具体来说,在流经导管的第二分支前,丝蛋白分子仍一直保持了先前的向列相液晶态;随着在第三分支内发生的离子交换过程和环境逐渐的酸化,丝蛋白分子链开始逐渐展开,而纺丝头施加的拉伸和剪切作用,使得蛛丝蛋白的分子链取向排列,并将其互相靠近,最终形成丝纤维。
丝蛋白分子处于大囊和导管前部时,通过采取液晶相的结构,使得丝蛋白链在导管内仍具有缓慢流动的能力。而在后续的流动过程中,分子链通过调整自身取向使之与流向保持一致,又可以一定程度上避免蛛丝蛋白被拉伸作用撕裂成众多小块,这有利于避免丝纤维内缺陷的产生,也有利于提高丝纤维的力学强度。从“液晶”模型中可以看到,拉伸和剪切作用对于丝纤维内的结构转变和丝纤维的力学性能起了重要的影响[27]。
至于“胶束”模型,则是基于蛛丝蛋白的一级结构,即,在蛛丝蛋白的氨基酸序列上,由于同时存在亲水性和疏水性的氨基酸残基,且这两种残基在分子链上交替出现,所以可以认为蛛丝蛋白具有一定程度的两亲性。正是由于亲水残基的存在,蛛丝蛋白在浓度较低时仍具有一定的溶解性[2]。而由于疏水基团的存在,蛛丝蛋白在水溶液中倾向于形成单分子链或多条分子链共同组成的胶束结构(类似表面活性剂)。随着蛛丝蛋白的分泌,丝蛋白溶液在导管内的浓度增加,越来越多的胶束开始形成并产生凝聚,共同构成了更大的球状结构。同样是在纺丝头,随着剪切和拉伸,这些球状结构沿着力的方向被拉长,同时,球状结构中的疏水部分被拉伸后开始相互靠近,通过这些疏水部分的疏水相互作用,更进一步增强了初生纤维的稳定性,最终导致了固化的丝纤维[2]。
显然,“液晶”模型和“胶束”模型最大的差别在于丝蛋白分子链在大囊腺体内的初始状态不同(分别是液晶态的向列相棒状聚集和胶束)。但需要指出的是,这两种模型并非完全相互矛盾。比如,若丝蛋白分子采取了所谓的“溶致液晶”相时,便仍可以形成规整的胶束结构[15]。
3.2 丝蛋白分子折叠:“成核-结晶”理论及丝蛋白材料晶体网络的介观重构
之前描述的丝蛋白分子“自组装机理”,尽管在一定程度上可以解释“纺体”中丝蛋白结构构象转变及丝纤维的纺出过程,但显而易见,这两种模型都十分看重拉伸和剪切的作用。在静置的丝蛋白溶液内,尽管缺失了剪切力,但仍能自发形成众多的纳米微纤,这是之前的理论不能解释的。考虑到微晶及晶体网络结构重要性,本课题组提出了丝蛋白折叠成更高级结构的另一种模型,即,丝蛋白分子的“成核-结晶”模型。这种理论基于晶体生长理论,认为丝蛋白分子首先通过连续的两个分子折叠过程,相继形成β折叠和β微晶,然后通过连续生长,建立后续的晶体网络。完整来看,丝蛋白材料主要结构的形成是由成核机制控制的。其中:第一步,丝蛋白分子通过成核完成由α螺旋和无规卷曲向β折叠的转变;第二步,众多β折叠通过互相堆叠(即结晶)形成β微晶;第三步,β微晶通过无定形链连接起来,形成具有纳米“渔网”状拓扑结构的微晶网络(即纳米微纤);第四步,众多的纳米微纤进一步生长和相互作用,构成纳米微纤网络。在分子折叠过程中,改变成核路径,调整成核动力学,就能改变丝蛋白材料在内的柔性材料的介观结构(介观重构),从而完全改变其性能[15]。
众所周知,结晶的热力学驱动力来源于系统自由能的降低。对应到丝蛋白结晶,由于初始的丝蛋白分子大部分呈无规卷曲或α螺旋构象,这样的丝蛋白分子保留了相当程度的柔性和可溶性,因而势能较高。随着丝蛋白分子的热运动或溶液内的环境发生变化,当邻近的丝蛋白分子距离缩小时(即局部浓度增加),稳定α螺旋结构的分子内氢键被破坏,当分子间氢键形成后,丝蛋白分子开始转变成β折叠结构。相比α螺旋和无规卷曲,β折叠的结构更稳定,且β折叠形成的速率正相关于丝蛋白分子的浓度,因此这样的构象转变过程实质上可以视为一个典型的成核过程。在第二阶段内,随着越来越多β折叠出现,β折叠之间的相互作用(如疏水相互作用、范德华力等“结晶力”)增强,促使来自不同分子的β折叠互相堆砌,导致了微晶的形成。在第三阶段中,当剪切力(如“纺体”中纺丝口的拉力)存在时,剪切力会导致第二阶段中形成的微晶沿着纤维轴向排列。当没有剪切力存在时(如丝蛋白溶液内),微晶的取向就倾向于无序。在第四阶段,由于不同形式的丝蛋白材料中,纳米微纤间相互作用的强度存在差异,且纳米微纤排列方式不同,导致丝纤维和其他丝蛋白材料的力学性能存在显著差异(见图6)[12,28]。以上的“成核-结晶”理论,不仅可以解释丝蛋白材料的晶体网络结构和介观多级结构的形成过程,并且能被用于指导丝蛋白材料的介观结构重构和实现功能化[15]。
为了更好地阐述丝蛋白分子的“成核-结晶”理论,在此首先介绍成核和结晶的基础知识。根据经典成核理论(CNT),只有当成核势垒被克服后,成核才会发生。此外,新形成的结晶核,只有在其尺寸超过临界尺寸后才在热力学上具有稳定性[13,29]。成核势垒ΔG*和临界晶核尺寸Rc的关系可以用公式(1)表示:
(1)
式中:ρc是核内的粒子密度;γ是区域表面自由能密度;Δμ是溶液相和结晶相之间的化学势差值。实际上,正是因为化学势存在差异,通过成核和结晶才能使整个体系的自由能降低。从公式(1)可以明显看出,Δμ是决定成核势垒和临界尺寸的主要因素,理论上,Δμ符合公式(2):
(2)
式中:a和C分别代表溶液中的活度和浓度;k是玻尔兹曼常数;T是温度;aeq和Ceq分别代表平衡活度和平衡浓度。以丝蛋白分子的成核和结晶为例,当改变丝蛋白分子的浓度,或改变丝蛋白分子的平衡浓度(这可以通过改变丝蛋白溶液的pH值、盐离子强度和温度等环境因素来实现),就可以改变成核势垒,从而影响丝蛋白的成核和结晶过程[13,29]。
3.2.1 调控热力学驱动力
具体来说,在丝素蛋白超分子材料的成核动力学中,主要需要关注的因素是蚕丝溶液浓度及温度。这两个因素共同组成了结晶成核的热力学驱动力:过饱和度。通常来说,成核速率J直接取决于其过饱和度,如公式(3)所示[13,29-30]:
(3)
式中:γcf代表结晶相和流动相的表面自由能密度;σ是过饱和度;k是玻尔兹曼常数;T是温度;B是常数。显然,过饱和度不仅与成核生长能垒密切相关,还会对成核速率、数目等起决定性的影响:体系的过饱和度较高时,结晶体系的成核速率及成核数目会增加,晶体结构的错配/晶界/缺陷也会相应的增加,整体体系最终有序度会下降,晶体网络也会趋于开放和多孔化。鉴于过饱和度对成核动力学的重要意义,通过调控合适的过饱和度,可以定向设计晶体结构及介观重构后续的更高级结构。具体来说,当丝蛋白分子浓度过低时,结晶所需要的时间过长,甚至不会有结晶发生。适当提高丝蛋白分子浓度可以有效增加结晶的可能性。当分子浓度过高时,丝蛋白分子结晶的速率过快,短时间内可以形成大量的晶核,这导致后续的晶体生长过程中由丝蛋白分子从溶液相向结晶相转变所需要的时间不足,将使丝蛋白分子更倾向于自组装成具有不规则结构的聚集体而非结构高度规则的β微晶。与改变丝蛋白分子浓度用以直接改变过饱和度的方法相比,当丝蛋白溶液内加入特定离子或者更改pH值时,往往会导致丝蛋白分子的溶解度发生变化(即所谓的“盐析”效应或分子带电),继而影响丝蛋白分子的过饱和度,同样会直接影响丝蛋白分子的结晶过程。
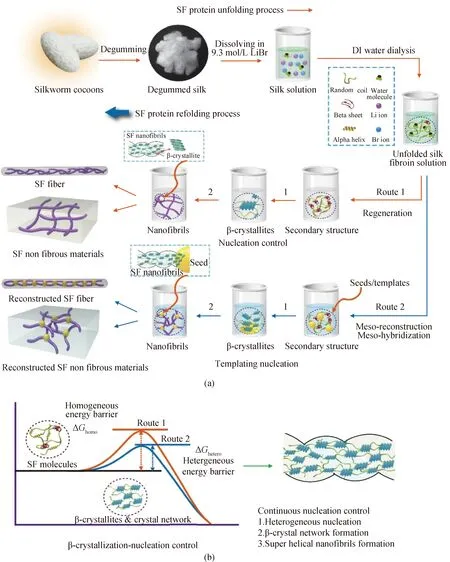
图6 丝蛋白材料的介观重构图 (a)丝蛋白分子的去折叠和重新折叠过程示意图,蚕茧经过脱丝胶、溶解、透析的过程后得到再生丝素蛋白溶液,再通过重新折叠,形成介观多级结构,其中,与性能最直接相关的关键结构因素包括微晶和晶体网络[28];(b)丝蛋白分子“成核-结晶”机理的示意图,丝蛋白分子的结晶必须克服所谓的成核势垒,当势垒较高时,可以通过异相成核的方式降低,促进丝蛋白分子结晶成微晶和微晶网络[12]Fig.6 Mesoscopic reconstruction of SF (a) Illustration of the SF protein unfolding process/refolding process, upon sericin removal, dissolve and dialysis, silk cocoons can be applied to prepare regenerated SF solutions, SF molecules in solution will spontaneously undertake refold/fold and form the mesoscopic hierarchical structures of silk materials, among different levels of structures, the key structural factors that mostly influence the macroscopic mechanical performance of SF materials are theβ-crystallites and crystal networks[28]; (b) scheme of the “nucleation-crystallization” mechanism, only after thenucleation barriers are overcome, the nucleation of SF molecules can occur, heterogeneous nucleation triggered by templating seeds can lower nucleation barrier, so as to promote SF molecules nucleation and formation of crystal network[12]
3.2.2 引进分子成核诱发剂(种子)促进异相成核
上述过程所描绘的场景中,丝蛋白分子发生成核的概率在整个系统中是处处均一的,这样的成核过程称之为均相成核。在实际的成核过程中,由于均相成核具有相当高的成核势垒,往往很难直接发生。在这样的情形下,考虑到体系中总是存在着异物或者基底(如杂质颗粒和盛放溶液的容器壁),发生成核的概率就无法处处均一。当这些异物或者基底能与溶液中的分子发生强烈的相互作用时,这些异物或基底便能显著地降低成核势垒;换句话说,在这些异物或基底的附近,发生成核的概率要远高于系统中其他部位。这种成核途径,被称为异相成核。对于丝蛋白材料,尽管它们本身已经具有优异的生物相容性和力学性能,然而单一组分的丝蛋白材料的结构可调性还是无法拓展。采用添加分子成核诱发剂(种子)诱导丝蛋白分子异相成核的方法,可以较大范围地调整蚕丝蛋白材料的成核动力学后续的介观结构重构,实现构建多组分复合的蚕丝蛋白超分子介观材料。
关于异物或基底对成核势垒降低的能力,可以用界面相关因子f(m)来量化。其中,参数m描述了晶体相与异物或基底之间的结构匹配程度。具体来说,若二者完美匹配,则认为此时的界面相关因子f(m)约等于0。这意味着,当成核相能在异物或基底上发生结构有序且定向的排列时,成核势垒会完全消失。与之相反,当结构匹配程度较差时(即f(m)趋近于1时),这样的异物或基底与成核相几乎没有任何结构上的相似性,在这种极端情况下,成核势垒等同于均相成核。在实际的异相成核过程中,界面相关因子的值通常在0到1之间浮动,这表明实际的初级成核过程总是在一定程度上受到异相成核的控制[13]。对蚕丝蛋白超分子材料的研究是一种极大的促进。如方程(4)所示:
(4)
式中:A是常数;f是描述异相成核相比均相成核时成核势垒下降的比例。只要介观成核模板的类型和其他条件保持不变,β微晶的成核速率J应与介观成核模板密度N0成正比[13,29]。此外,由于添加剂分子会参与到丝蛋白β微晶的成核速过程,不同的添加剂分子与蚕丝蛋白的相互作用强度会有所差异,同时不同添加剂分子的分子大小及结构强度也不相同,而这些对蚕丝蛋白材料中β微晶的成核及结构都会有很大的影响。因此,当添加具有不同特性的添加剂时,理论上可以制备出相应的蚕丝蛋白复合超分子材料,以期获得一定程度上具有添加剂分子性质的功能化丝蛋白材料。迄今,已有多种种子,包括(但不限于)角蛋白、BSA、BMP、PAA、PSS等被用于分子成核诱发剂[31]。例如,Chen等[31]已证实,单分散的表面修饰有功能化羧基的聚苯乙烯(PS)纳米颗粒可以作为一种理想的异物,诱导丝素蛋白在其表面发生异相成核。具体来说,由于这些纳米颗粒表面的羧基可以通过和丝素蛋白肽链上的氨基发生氢键作用,能降低丝素蛋白的成核势垒,从而加快丝蛋白分子成核,促进β折叠和β微晶网络的形成。这已经分别通过激光共聚焦显微镜和扫描电镜实验进行了证实。而Ma等也证实,单分散的碳纳米管(CNTs)也可以作为一种有效的分子成核诱发剂(见图7)[32]。值得一提的是,这样得到的复合蚕丝纤维,不仅由于介观结构重构改善了力学性能,还由于碳纳米管参与到蚕丝纤维的结构中,赋予了复合材料一定的导电性,为该复合材料应用在后续的柔性电子器件中提供了可能[11]。
除了聚苯乙烯和碳纳米管,石墨烯也被证实能对丝蛋白分子的成核与结晶过程产生影响。Ling等[33]证实,当石墨烯片层加入丝素蛋白水溶液并浸泡一段时间后,纳米微纤会在石墨烯片层的表面发生选择性极强的聚集。而在石墨烯片层没有覆盖的基底上,则鲜有纳米微纤的形成与分布(约1%)。这种富集作用,本课题组认为是来源于疏水性的石墨烯片层与丝蛋白分子间的强烈疏水相互作用。
以上通过引进分子成核诱发剂(种子)促进异相成核来调控丝蛋白分子的结晶动力学,必然导致后续丝蛋白材料的介观结构重构,例如改变材料的结晶度和晶体密度即取向度等,最终实现丝蛋白材料的宏观性能改变。比如,Chen等[31]的实验结果证明,添加聚苯乙烯小球后的复合纤维的断裂强度和弹性模量比单纯组分的再生蚕丝纤维几乎翻了一倍,且当添加的聚苯乙烯小球浓度控制在一定的范畴内时,制得的复合纤维的断裂强度会随着添加物浓度增加而升高。至于多壁碳纳米管对复合丝蛋白材料性能的影响,也同样被证实,当添加的碳纳米管的质量分数小于1%时,可以显著提高复合蚕丝纤维的韧性。而当碳纳米管浓度进一步增加后,复合纤维的性能反而变差。值得注意的是,并非所有的分子成核诱发剂都有利于丝蛋白分子的异相成核,当添加物本身具有高度的自组装倾向时,这样的添加物在加入纺丝原液后会快速形成添加物团簇,而这样的团簇会成为复合纤维内的缺陷,导致复合纤维的力学性能下降。对于这样的添加物来说,加入合适的表面活性剂使添加物在纺丝原液内均匀分散就显得特别重要。如前所述,这种通过添加特定添加物制备复合人工纺丝纤维还具有另一个优势:由于特定添加物可能具有自身独特的性能,当添加物被掺杂进复合人工纺丝纤维的多级结构中后,还很有可能实现介观掺杂,即制备功能化的丝蛋白材料[34]。
3.2.3 介观掺杂功能化
基于柔性材料多级结构的形成,主要是由成核控制这一事实, 在柔性材料折叠过程中,加入功能性纳米“种子”,能使其嵌入到柔性材料的介观结构中,使其具备原先单一组分的柔性材料不具备的一些功能[11]。介观功能化的概念如下:将功能化分子或功能纳米粒子组装到丝蛋白材料的介观网络中,使丝蛋白材料在保持原有特性的基础上,被赋予全新的功能,实现复合丝蛋白材料的介观功能化(见图8)[34]。通过介观功能化,已成功制备出具有新性能的丝蛋白材料,如发光、导电、发荧光、磁性等[8,10-11,33,35-39]。

图7 (a)添加不同聚苯乙烯小球后的复合丝素蛋白溶液的溶胶-凝胶转变过程;(b)添加了不同聚苯乙烯小球后的复合丝素蛋白溶液的光密度变化过程;(c)成核速率与添加的分子成核引发剂浓度N0的线性关系;(d)模拟显示丝蛋白分子倾向于聚集在碳纳米管周围;(e)模拟的丝蛋白-碳纳米管复合体;(f)将聚苯乙烯纳米颗粒浸入丝蛋白溶液30 min和120 min后,利用扫描电镜对其形貌进行表征,能清楚观察到直径约为30 nm的纳米微纤出现在颗粒表面;(g)丝蛋白-碳纳米管复合丝蛋白材料的介观结构重构的示意图;(h)通过原子力显微镜观察到的丝蛋白纳米微纤网络;(i)通过原子力显微镜观察到的丝蛋白纳米微纤-碳纳米管复合网络[11,32]Fig.7 (a) Storage moduli of SF-PS mixtures at various PS concentrations as a function of time; (b) variation in the normalizedoptical density (OD) of neat SF solution and SF-PS mixtures with different PS concentrations; (c) linear correlation betweennucleation rate J and density of the added nucleation seeds N0; (d) representative all-atom simulation snapshots of the initial SF-CNF structure (five silk biomolecules) and those after relaxation (last structure). Blue and orange colors represent CNF and SF, respectively; (e) morphology of the SF-CNF assembly; (f) scanning electron microscopy (SEM) images of thePS particles incubated in the SF solutions for different periods; (g) schematic illustrations of the mesoscopic network reconstruction(SF-CNTs); (h) atomic force microscopy (AFM) image of SF nanofibrils networks; (i) AFM image of CNTs/SF hybrid mesoscopic networks[11,32]

图8 (a)丝蛋白柔性材料功能化和纳米种子与丝蛋白分子的相互作用协同示意图;(b)通过种子对丝蛋白分子折叠过程中的诱导成核,不同功能性纳米种子,如金纳米簇、银纳米簇、碳纳米管、上转换粒子、石墨烯等,都可以被掺杂到柔性材料的介观结构中,实现介观柔性材料的功能化[33-34,38-40]Fig.8 (a) Schematic of mesoscopic SF functionalization and interaction synergy of nano seeds with SF; (b) brief summary of nano seeds (e.g., gold nanoclusters, silver nanoclusters, carbon nanotubes, upconverting nanoparticles, graphene) used to functionalize SF and the potentially enhanced properties[33-34,38-40]
除了上文提到的通过碳纳米管制备导电丝蛋白材料,ZnSe、CdTe等量子点也可以作为功能化组分,通过分子识别功能化,成功组装到蚕丝纤维和丝蛋白薄膜中,制备可以发出白光的蚕丝膜光学器件(见图9)[10]。类似地,将上转换纳米粒子组装到丝蛋白支架的介观结构后,通过红外激光,能实现深入体内的无线充电、信息交换与近红外医学成像(见图10)[10,28,34,40]。

图9 量子点功能化电致发光蚕丝膜及器件[10](a)各种类的量子点在302 nm激发光下的荧光光谱,插图为相应的光学照片;(b)量子点与丝蛋白复合膜及器件的电致发光能力稳定;(c)显示量子点丝蛋白复合膜的电致发光能力的照片Fig.9 Quantum dots incorporated into mesoscopic SF membranes to synthesize functional electroluminescence devices[10](a) characterization of different quantum dots which were embeded into composite SF membranes; (b) SF/QDs composite membranes display stable electroluminescence ability; (c) image showing electroluminescence ability of the SF/QDs composite membranes and devices

图10 丝介观光子晶体与上转换粒子介观功能化结合组成介观光子器件,通过红外激光,可以实现对生物体内植入器件深入体内的无限充电、信息交换与控制[10]Fig.10 By functionalize mesoscopic silk photonic crystals with upconverting nanoparticles (UCNPs), mesoscopic functional photonics devices can be synthesized and been applied in areas of implantable medical devices[10]

图11 利用介观功能化策略所构建的银纳米簇掺杂丝蛋白基柔性忆阻器[12,37]。(a)相关的TEM照片,展示嵌入到丝蛋白材料结构中的银纳米簇尺寸;(b)银纳米簇掺杂丝蛋白材料的原子力显微镜高度图(左)和表面开尔文电势分布图(右)。电子阱(纳米环区域)由于银纳米簇的存在,容易导致电子的积累,呈现较低的电势;(c)电势分布;(d)经典密度泛函理论Poisson-Nernest-Planck模拟Fig.11 Silver nanoclusters doped silk fibroin based flexible memristor constructed by mesoscopic functionalization strategy[12,37](a) TEM image of functionalized SF-based memristor; (b) AFM image and Kelvin probe force microscopy (KPFM) images of AgNC (white circles)-functionalized SF film, the electronic wells are 200 nm in size and have significantly lowelectric potentials due to the accumulation of electrons at the surface; (c) corresponding electronic potential; (d) simulation ofclassical density functional theory using the Poisson-Nernst-Planck model
至于如纳米金簇、银簇介观功能化的蚕丝材料,由于其介观电子结构与本征蚕丝材料完全不同,还可以将之用于构建全新的、性能优异的柔性介观忆阻器。最近,利用基于丝蛋白的柔性介观忆阻器及人工神经突触元器件方面,已经取得了突破性的进展(见图11)[12,37]。与同类有机生物元器件相比,其速度是有机生物材料的上百倍,耗电只是最好的同类有机生物电子器件的十分之一,开关比达到1 000,且重复性、稳定性十分优异。该特性可在电场作用下实现对材料忆阻性能的优化调控,得到擦写速度达10 ns的超快蛋白质基阻变存储器,比纯蚕丝以及目前所报道的蛋白质、有机基忆阻器快二到三个数量级。如此快的擦写速度,甚至可以与顶级无机类材料忆阻器相媲美[12]。这证实了介观功能化策略在柔性材料功能化领域具备极大的推广价值,可为柔性电子器件提供新的设计思路及理论基础。同时,对未来实现可植入生物电子传感与计算、在实时人工智能计算、远程人性化人工智能医疗等,具有开创性意义。值得一提的是,基于导电丝蛋白材料的柔性电子器件,还已经被证实可以用于发展各种性能优异的蚕丝柔性介观传感器,包括蚕丝蛋白基柔性压力传感器、拉伸传感器、温度传感器、湿度传感器、生化传感器(pH传感器、血糖传感器、尿酸/乳酸传感器)等[12,41]。
4 晶体生长理论在其他丝蛋白材料中的应用
由于丝蛋白分子良好的生物相容性、可控的降解速率及独特的光学和电学性能,利用丝蛋白分子构建新一代的绿色生物材料器件具有非常大的前景。人类利用天然蚕丝纤维已有数千年历史,近年来,越来越多的学者也开始对丝纤维介观结构与宏观力学性能之间的关系进行研究。这些研究不仅深化了对丝纤维的认识,获得的知识和经验还指导了科研人员如何制备性能优异的非纤维状丝蛋白材料,如丝蛋白薄膜,水凝胶及支架材料。与上文中制备力学性能提升的丝纤维的方法类似,在制备非纤维状的丝蛋白材料时,同样也可以利用晶体生长理论作为指导,在介观尺度上对其多级结构特别是晶体和晶体网络进行重组,最终实现其力学性能的提升和功能化。在本小节中,将以丝蛋白薄膜为例,介绍晶体生长理论在其中的应用。利用丝蛋白水溶液制备丝蛋白薄膜时,传统的方法包括浇筑法和旋涂法。这两种方法的基本原理都是设法将溶剂分子从丝蛋白溶液中去除,从而增加丝蛋白分子的局部浓度(即过饱和度),最终引起相分离和结晶(即成膜)。首先以浇筑法为例阐述丝蛋白薄膜的成膜过程。在丝蛋白溶液中,丝蛋白分子仍处于无规卷曲或α螺旋构象,所以此时的丝素蛋白分子仍然是可溶。当一定体积的丝蛋白溶液被浇筑在不同的基底后,伴随着水分子的逐渐蒸发,丝素蛋白分子的浓度逐渐升高,考虑到之前提过的“成核-结晶”机理,显然,这样的过饱和度增加可以引发素蛋白分子的结晶,最终形成丝蛋白薄膜。而在旋涂法中,除了溶剂自发蒸发过程带来的过饱和度增加,在溶液被旋转过程中产生的剪切力,也同样对丝素蛋白分子的结晶动力学产生重要影响。
以上两种方法虽然简单易行,但仍具有一定的局限性。浇筑法制备的丝蛋白薄膜常常产生皱缩现象。而对于旋涂法制备的丝蛋白薄膜,又常常由于剪切力的分布,导致薄膜边缘的厚度高于中央。近年来,为了制备厚度可控、形貌理想、具有特殊用途的超薄或特种丝蛋白薄膜,学界和工业界投入了大量精力并提出了以下几种方法。
Bressner等[42]报道了一种所谓的“电凝胶成膜”技术,该技术使用具有特殊闭环结构的阳极作为模板,通过施加电场来促进丝素蛋白分子的溶胶-凝胶转变,可以短时间内得到形状由阳极决定的丝蛋白凝胶。随后,将该凝胶进行干燥处理,便可以获得透明的丝蛋白薄膜。显然,这种方法最大的优点是可以很方便地制作特定形状的丝蛋白薄膜。
Jiang和Tsukruk等[43]开发了一种先进的“旋涂辅助多层沉积”(spin-assist layer-by-layer, SA-LBL)的技术来实现对丝蛋白薄膜厚度的精确控制。这种技术可以通过控制旋涂的次数来精确地调节丝蛋白膜的厚度。基于类似的思路,Wang等[44]也开发了一种“逐步沉积”的方法,制备了厚度可控的多层丝蛋白膜。
与丝纤维材料类似,丝蛋白薄膜的宏观力学性能,也在很大程度上取决于内部的介观结构。举例说,丝蛋白薄膜的结晶度便与它的力学稳定性和降解率直接相关。薄膜内二级结构(特别是α螺旋)的含量,则与薄膜的透气性和透水性直接相关。为了提高丝蛋白薄膜的稳定性和模量,迄今已存在不少方法,如,将丝蛋白薄膜浸渍入甲醇或乙醇中,水蒸气退火处理[45],将丝素蛋白溶液与其他高分子进行掺杂后制备复合丝蛋白薄膜[45],将制备的初生丝蛋白薄膜进行单轴后拉伸等(见图12)[15,46]。上述方法实质上是对丝蛋白薄膜内部的多级结构进行了介观重构,而且,这些方法都符合本课题组提出的丝蛋白分子“成核-结晶”理论。

图12 (a)对丝蛋白薄膜进行乙醇处理后,β折叠的含量上升[47];(b)将不同比例的丝蛋白分子和羊毛角蛋白分子进行混合,制备的复合蛋白薄膜的力学性能会随着比例而变化[45];(c)对丝蛋白薄膜进行单轴后拉伸,内部微晶的取向变好,抗断裂能力随之增强[15,46]Fig.12 (a) Inuence of ethanol treatment on β-sheet content, the incorporation of ethanol would increase content of β-sheet[47]; (b) mechanical performance of SF/WK composite films can be altered by changing the ratios of WK[45];(c) after extension, the orientation function is improved so that the breaking strength of SF films become larger[15,46]
与初生丝蛋白纤维相似,初生的丝蛋白薄膜由于内部的结晶度较低,所以力学性能较差。此时可以采用甲醇或乙醇浸渍的方法,促进丝蛋白分子从无规卷曲或α螺旋向β折叠的构型转变,从而提高丝蛋白薄膜的结晶度和力学性能。至于单轴后拉伸对丝蛋白薄膜结构的影响,则与人工纺丝中对丝纤维进行后拉伸的处理类似。单轴后拉伸前后的丝蛋白薄膜的WAXS光谱清楚显示,单轴后拉伸可以显著提高薄膜内的丝蛋白分子链的取向度(使之沿着单轴拉伸方向重新排列),相应地,薄膜的机械性能得到显著增强。除了单轴后拉伸,制作丝蛋白薄膜的原液中丝蛋白分子的浓度、金属离子的种类、制膜时的基底的选择等因素,也会对丝蛋白薄膜的介观结构和宏观力学性能产生影响。具体来说,当丝蛋白溶液的浓度增加时,获得丝蛋白薄膜的速率会加快。当丝蛋白溶液的浓度在一定范围内时,薄膜的厚度还与加入溶液中的金属离子(如钠离子)的浓度成正比。这是因为金属离子加入后可以提高丝蛋白分子间静电相互吸引作用,因此有利于结晶(即成膜)。至于基底的影响,现已证实丝蛋白分子更容易在疏水基底上发生成膜。这是因为疏水基底更容易与丝蛋白分子发生疏水相互作用,从而促进丝蛋白分子在这样的基底上发生成核和结晶行为。与复合丝蛋白纤维一样,将丝蛋白溶液中混入特定的高分子聚合物(如羊毛角蛋白等),也可以制备性能可调控的复合丝蛋白薄膜[45]。不仅如此,改变丝蛋白和羊毛角蛋白的比例,还可以方便地调节复合薄膜的弹性和可降解能力。具体来说,当丝蛋白比例较高时,会导致复合薄膜内的β折叠含量增多,因此有利于增加其韧性,减少其降解能力。与之相反,当羊毛角蛋白含量较多时,α螺旋含量会增多,这样的复合蛋白薄膜柔性和弹性更好,且更容易从形变中恢复[45]。
5 结语与展望
本文对柔性材料特别是丝蛋白材料的晶体和晶体网络结构进行了深入而详细的阐述,重点讨论了丝蛋白分子折叠过程中涉及的“成核-结晶”机理,并证实了该机理的正确性。在该机理的指导下,还拓展了多种控制丝蛋白分子成核与结晶的技术手段,相应地对丝蛋白材料的结构进行介观重组,最终实现了性能的改善和介观掺杂功能化。这种思路为未来创造更强的高性能软物质材料提供了理论指导,并指明了发展方向。

刘向阳,教授,国家特聘教授,教育部长江学者,博士生导师,厦门大学生物仿生及软物质研究院院长。发表SCI论文400多篇,其中顶级杂志>90篇(Nature3篇),出版英文专著6部。主要研究领域为晶体生长、仿生材料、柔性材料及相关功能化、柔性电子、柔性传感器件和能源器件,表面以及胶体科学。在国际国内学术会议及研讨会上作大会报告及邀请报告150余次,主办国际会议30余场。担任亚洲晶体生长和晶体技术协会第五届主席,国际晶体生长协会理事,AdvancedFunctionalMaterials(Wiley, 2016)、Small(Wiley, 2015)、SmallMethods(Wiley, 2020—2021)、《物理学报》(编辑,2016—)等杂志专刊客座主编。
猜你喜欢
学与玩(2022年12期)2023-01-11
人工晶体学报(2021年10期)2021-11-26
陶瓷学报(2021年4期)2021-10-14
建材发展导向(2021年11期)2021-07-28
陶瓷学报(2021年1期)2021-04-13
制造技术与机床(2017年4期)2017-06-22
中国塑料(2016年9期)2016-06-13
中国塑料(2015年7期)2015-10-14
火炸药学报(2014年1期)2014-03-20
