
硬粒小麦ANW16F株高及构成因子与部分产量性状遗传研究
2021-05-25 03:30龚胤书魏淑红彭正松杨在君钟明志张家敏
西南农业学报 2021年2期
龚胤书,魏淑红*,彭正松,杨在君,钟明志,张家敏
(1.西华师范大学生命科学学院,西南野生动植物资源保护教育部重点实验室,四川 南充 637009;2.西昌学院农业科学学院,四川 西昌 615013)
【研究意义】Rht(Reduced height) 是小麦矮化育种的关键基因。迄今为止,已发现并正式命名编号的Rht基因有 25个[1]。大多数Rht基因具有较强的抗倒伏性和肥水反应,但存在一些缺点,限制了在生产上的利用[2]。如Rht4、Rht5、Rht6、Rht7在降秆同时,大大降低了小麦产量[3]。Rht3、Rht10、Rht12以及Rht21降秆作用太强,导致粮食产量过低[4]。目前推广小麦品种中被广泛应用的Rht基因主要是Rht-1、Rht-2和Rht8,Rht9亦有局限使用[5-6],可见小麦育种实践中矮源十分单一。此外,对矮秆基因的研究主要集中在六倍体小麦,对四倍体小麦矮秆基因研究较少,育种利用也较局限。因此,发掘新矮源,寻找具有育种潜力的矮秆基因,对小麦高产及抗性育种十分重要。ANW16F 是携带显性矮秆基因Rht16的硬粒小麦(Triticumdurum,2n=4X=28,AABB)[7]。对硬粒小麦ANW16F株高与构成因子及产量性状进行遗传研究,将为进一步利用Rht16基因奠定基础。【前人研究进展】大约公元前8000年,中亚地区祖先在两河流域的肥沃土地上开始种植小麦等作物[8]。小麦作为中国乃至全世界最重要的粮食作物之一,高产一直是育种学家追求的首要目标。较高、较细的茎秆容易发生倒伏、发生病害等,降低小麦产量[9],因此提高抗倒伏能力是小麦高产的重要途径[10]。矮秆小麦由于抗倒伏能力强,适宜高度密植,群体光能利用效率高,大大提高了产量。20世纪60年代中期,利用日本“农林10号”(Norn10,含矮秆基因Rht-1和Rht-2)小麦品系育成了30多个矮秆、半矮秆品种,这些品种同时具有抗倒伏、抗锈病、高产的突出优点,在发展中国家迅速推广,产生了巨大效益,被称为“绿色革命”[11]。目前发展中国家种植小麦1亿公顷,其中60%都是“绿色革命”的育种成果。“绿色革命”已经证明矮秆基因对提高小麦的产量的巨大作用。硬粒小麦是四倍体小麦中唯一仍在种植的物种,具有品质好、籽粒蛋白质含量高、抗性强的优点。硬粒小麦具有与普通小麦(TriticumaestivumL.,3n=6X=42,AABBDD)同源的A、B基因组,易于种间杂交实现染色体配对、易位和重组,达到利用有益基因改良普通小麦的目的[12-13]。前期研究表明,ANW16F 对外源赤霉素反应敏感,矮秆性状稳定,降低株高明显,分蘖多,叶片宽,茎秆粗壮[14-15]。也有报道认为Rht16在高山较阴凉的环境下不能抽穗[16]。采用 SSR标记将Rht16基因初步定位于染色体 6AS上,位于SSR标记Xbarc3和Xgwm356之间,距离Xbarc3和Xgwm356均较远[17]。由上述研究进展可知,目前对Rht16的研究利用非常薄弱。ANW16F株高遗传特点,株高与构成因子及产量性状的相关性、株高及构成因子的数量遗传模型等尚不明确,有待研究。【本研究切入点】本研究利用矮秆硬粒小麦ANW16F和高秆LD222以及杂交F1代、F2代群体,对株高及其构成因素、部分产量性状等10个农艺性状进行表型特征和相关性分析,并利用单个分离世代的群体主基因+多基因混合模型方法进行遗传分析。【拟解决的关键问题】此研究结果以期为Rht16基因的育种利用及后续利用分子标记定位主效QTL提供参考。
1 材料与方法
1.1 试验材料
以矮秆硬粒小麦材料ANW16F为母本,高秆对照硬粒小麦材料LD222为父本杂交获得F1代,F1自交得F2代群体共284株。ANW16F和LD222由日本岐阜大学农业学院Watanabe提供[15]。试验在西华师范大学生命科学学院实验田进行,播种前施氮肥、磷肥,田间种植及管理均按当地大田生产模式,播前灌溉以保证出苗。
1.2 性状测定
于成熟期对亲本、F1以及F2代植株进行室内考种。测量株高、穗下茎节长、倒2至倒5茎节长、穗长、单株穗数、有效穗数、小穗数。有效穗数为穗粒数超过5粒的小麦穗数。
1.3 数据分析与统计
数据导入Excel表,由origin 8.0及Rstudio分析绘图得到图表,进行相关性分析和方差分析。参考王建康、盖钧镒、章元明等提出的主基因+多基因遗传混合模型利用Rstudio对ANW16F/LD222杂交F2后代共284株植株的性状,包括株高及各个节间长进行遗传分析[18-20]。矮秆基因的相对效应计算公式:(矮秆平均株高-高秆平均株高)/高秆平均株高×100 %[21]。显著性差异:*P<0.05,**P<0.01,***P<0.001和****P<0.0001。
2 结果与分析
2.1 株高、穗长、节间长度表型特征分析
矮秆亲本ANW16F平均株高103.61 cm,高秆亲本LD222平均株高157.77 cm。ANW16F相较于LD222降低株高约33.9 %(P<0.05)。F1代株高均值为119.97 cm,与矮秆亲本较为接近。F2代株高平均值为105.01 cm。F2代有高秆和矮秆分离,暗示矮秆是显性性状。F2代呈双峰且明显偏矮秆性状的偏态性分布,且偏度值和偏度值都小于3,表明ANW16F株高性状受主基因控制(图1)。
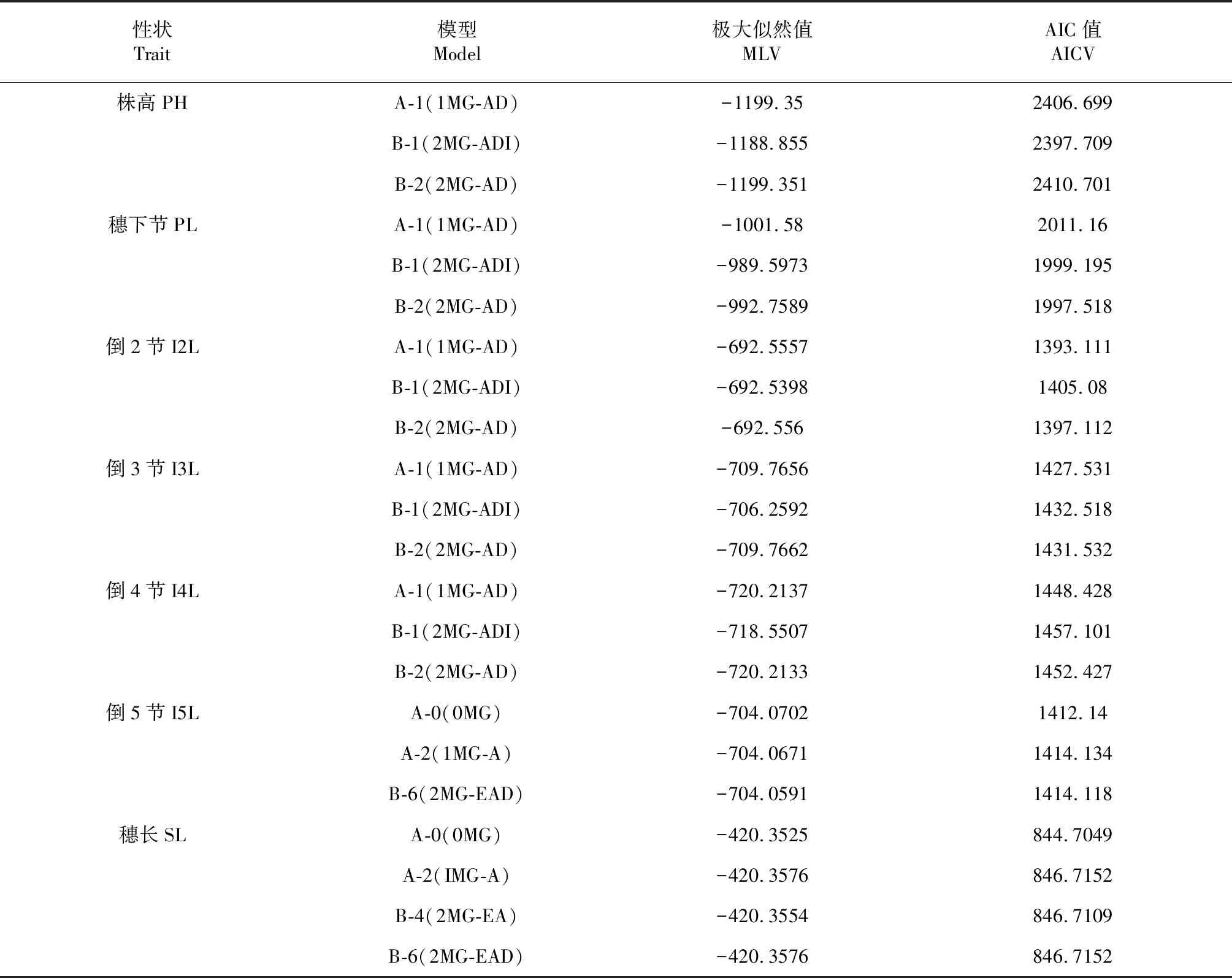
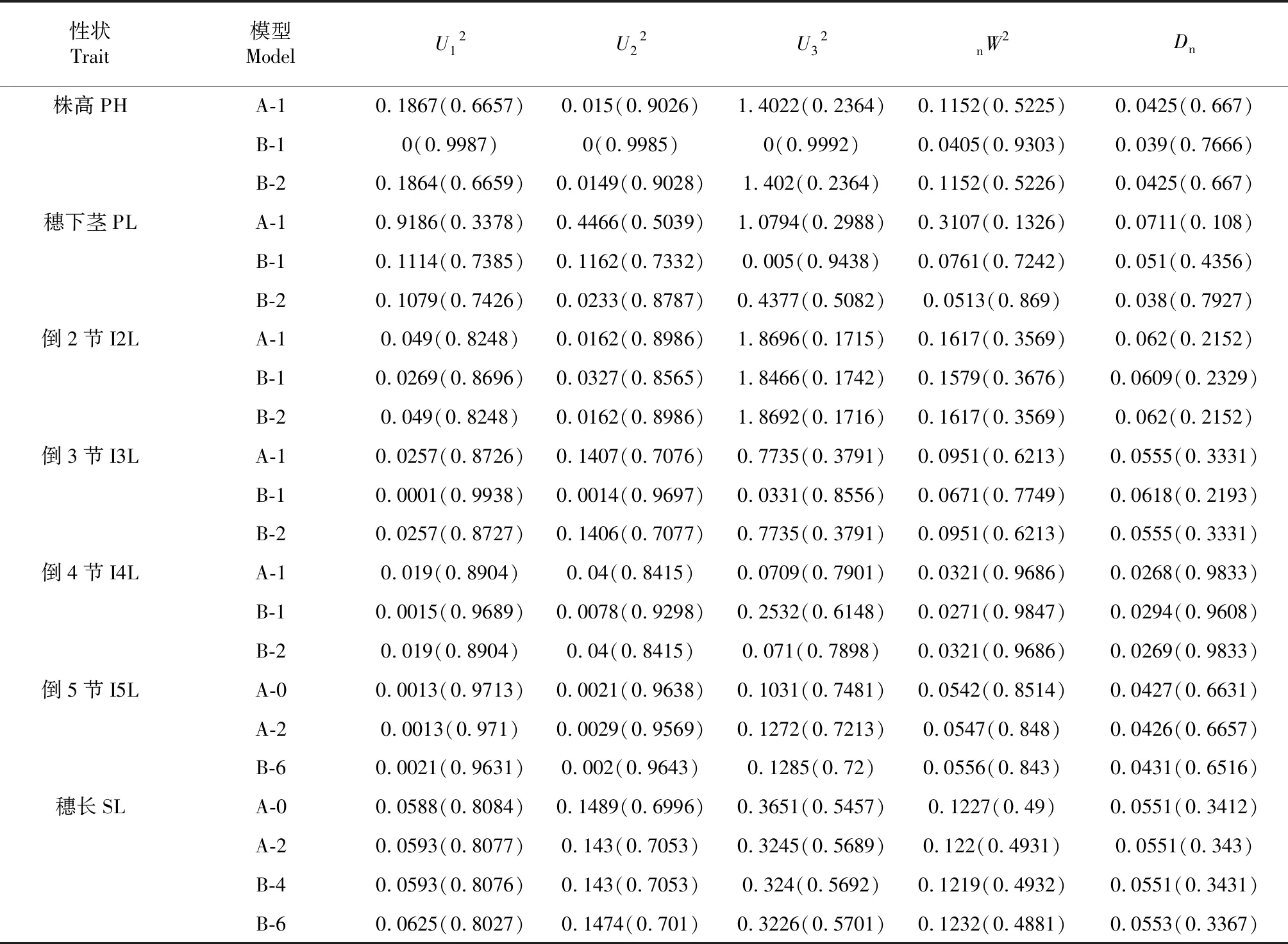
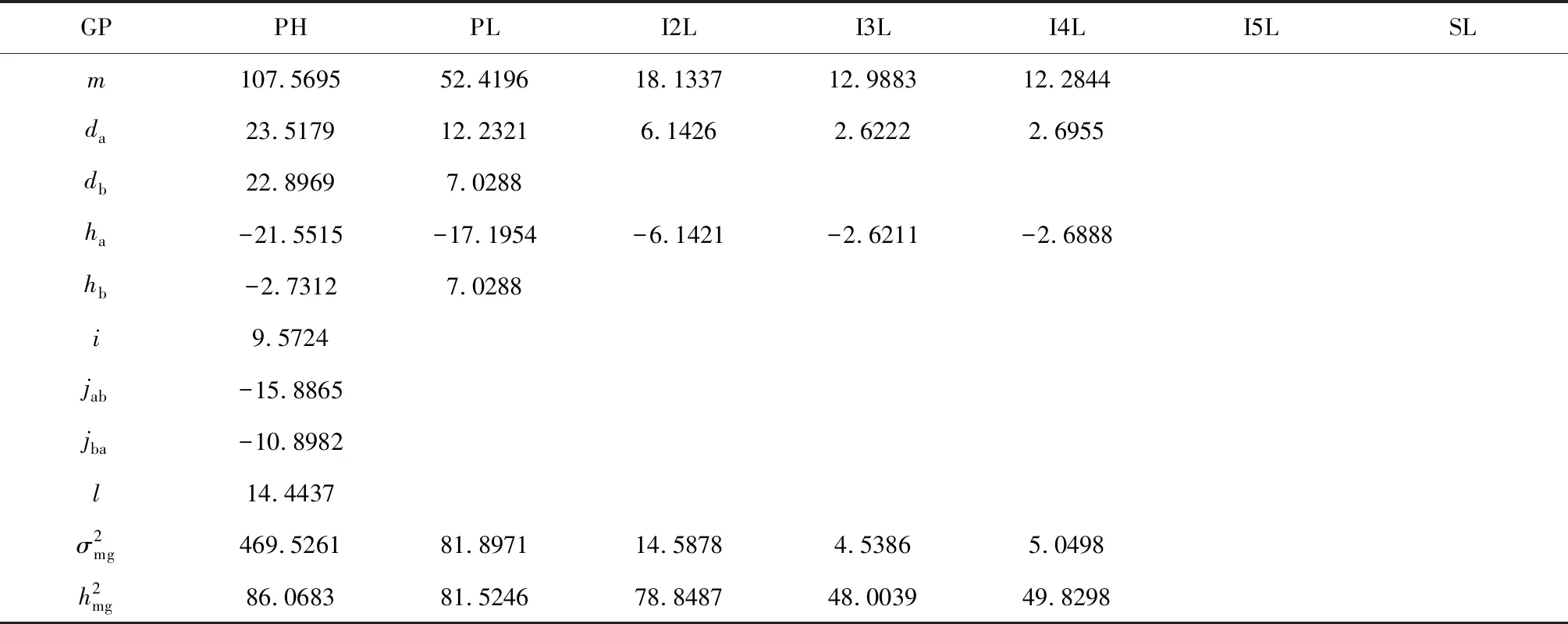
株高由穗长与各节间长度共同构成。除了穗长外,矮秆ANW16F各节间长均比高秆对照LD222短(P<0.05),各个茎节缩短能力依次PL 用R语言可视化Corrplot包和Performance Analytics包进行相关性和显著性分析。由图3得知,株高与各茎节均为极显著正相关,其中与倒2节相关性最高,其次为穗下节。株高与小穗数、穗长与小穗数为显著正相关,且穗长与小穗数相关性高于株高与小穗数的相关性。穗数与有效穗数相关性最高,达到0.98。 2.3.1 最适遗传模型的建立 参考盖钧镒等方法[22],对ANW16F/LD222 F2代的株高、穗长、茎节长进行数量遗传分析。选择AIC值最低及与其接近的3个模型为备选模型,获得极大似然值与AIC值(表1)[23]。 进行均匀性检验、Smirnov检测、Kolmogorov检验获得F2性状最优模型和备选模型的5个参数:U12、U22、U32、nW2、Dn,并进行筛选,最终选择统计量达到显著水平个数较少的模型作为最优模型(表2)。株高的3个备选模型的5个统计量的P值均不显著(P>0.05),模型B-1的AIC值较A-1、B-2最小,因此确定B-1遗传模型为株高的最优模型,表现为由两对加性-显性-上位性主基因控制的遗传模型。穗下茎节的最优遗传模型为B-2,表现为两对加性-显性主基因遗传模型。倒2、倒3、倒4节间的最优模型为A-1,表现为一对加性-显性主基因遗传模型。倒5茎节和穗长的最优模型为A-0,不受主效基因控制的遗传模型。 2.3.2 遗传参数的估计 根据迭代ECM算法,即IECM算法,利用筛选出的最适遗传模型,估计F2代株高、穗长、茎节长的遗传参数,包括一阶遗传参数与二阶遗传参数[24]。 表3显示,株高,da、db均为正值,且较为接近,说明2对主基因增效接近,|ha/da|<1,|hb/db|<1,说明2对主基因均为部分显性,以加性效应为主。ha和hb均为负值,且|ha|>|hb|,说明2对主基因都是矮秆对高秆为部分显性,且第1对主基因的部分显性效应大于第2对主基因的部分显性效应。|jba+jab|>|l|>|i|,说明2对主基因间存在互作效应。株高主基因的遗传率为86.07 %,说明株高受遗传因素影响较小。 穗下节,da、db均为正值,且da>db,说明第1对主基因增效大于第2对。|ha/da|>1,|hb/db|=1,说明2对主基因均为完全显性,且第1对主基因以显性效应为主,第2对主基因显性效应和加性效应同等重要。ha为负值,说明第1对主基因短穗下节对长穗下节为显性,hb为正值,说明第2对主基因长穗下节对短穗下节为显性。|ha|>|hb|,说明第1对主基因的显性效应大于第2对主基因的显性效应。主基因的遗传率为81.52 %,说明穗下节长受环境影响较小。 表1 ANW16F/LD222 F2代株高、穗长、茎节长极大似然值与AIC值 表2 遗传模型的适合度测验 倒2、3、4节均为ha<0,且|da|≈|ha|,说明短茎节相对于长茎节为部分显性,且显性效应和加性效应同等重要。倒2节长遗传率为78.85 %,倒3、4节长的遗传率较低,分别为48 %、49.83 %,说明倒2节受环境影响较小,而倒3和4节受环境影响相对较大。 小麦株高由穗长和地上部分各节间伸长构成。大量研究发现,矮秆基因能够显著缩短穗长和各节间长,或者只是显著缩短节间长,而穗长变异不大。比如Rht14通过缩短各节间长度降低株高27 cm,降秆能力约25 %。Rht18由于穗和节间远端缓慢生长,导致普通小麦降低株高最高可达35.0 %[25]。王健等[26]研究也发现,30个冬小麦矮秆品种株高降低是由于各节间长缩短所致,尤其是倒2、3茎节。崔淑佳等[27]利用20个高、中、矮秆小麦品种研究同样证实株高降低与节间长缩短有关,其中穗下节在株高构成中贡献较大,穗长在高、中、矮小麦中变化系数不大。在本研究中,矮秆小麦ANW16F与对照高秆小麦LD222相比,降秆33.9 %,穗长对株高变异影响不大,各节长差异均为显著(P<0.05),各茎节缩短程度为PL 株高与其构成因素、产量性状相关性研究较多。多数学者认为株高与其构成因素以及产量性状正相关[28-30]。但朱新开等研究表明,在一定株高范围内,株高及茎节长(除穗下节)与产量性状呈负相关[31]。本研究中,株高与其构成因素、产量性状均呈极显著或显著正相关,其中与穗下节、倒2节长相关性较高,且株高与各节长的相关性高于株高与穗长的相关性,相邻节间长的相关性也较高。 表3 F2代株高、茎节长及部分产量性状遗传参数估计值 盖钧镒等优化传统数量性状多基因遗传模型,并初步建立植物数量性状遗传体系分离分析方法,依据7类模型,即A-1对主基因、B-2对主基因、C-多基因、D-1对主基因加多对主基因、E-2对主基因加多基因、F-3对主基因、G-3对主基因加多基因,与主基因加多基因的显性、上位性、加性效应组合构成多种模型,针对不同世代的性状表型进行遗传模型评估[32]。在P1、P2、F1群体、F2群体、回交世代等不同群体均可进行此数量性状分离分析方法[19-20,33]。目前广泛用于多种植物,如水稻[34-35]、油菜[36]、棉花[37]、玉米[38]、小麦[39]等。小麦株高及其构成因素等主基因+多基因混合遗传模型研究较多,或为一对主基因+多基因控制模型[40-41],或为两对主基因+多基因混合模型[42-43],或为无主基因而是多基因控制模型[39-44]。在遗传效应方面,主要是加性效应+显性效应,且以加性效应为主,上位性效应较少[40-41,43,45-46]。本研究中,株高表现为由两对加性-显性-上位性主基因控制的遗传模型,且两对主基因均以加性效应为主,穗下节长由两对加性-显性且以显性效应为主的主基因控制,倒2、3、4节长均为由一对显性效应和加性效应同等重要的主基因的控制。倒5节长和穗长不受主基因控制。株高与株高构成因素遗传模型与前人研究略有不同,这与研究材料遗传背景和所处的环境不同有关。株高、穗下节长、倒2节长主基因的遗传率较高,依次为86.07 % 、81.52 %和78.85 %,说明受环境影响较小,倒3、4节长的遗传率较低,仅为48 %和49.83 %,受环境影响相对较大。 综合考虑株高构成比例、相关性分析,以及遗传率的大小,今后在利用Rht16基因进行矮化高产育种时,穗下节和倒2节可作为重要的育种筛选指标,且可在较早期选择。本研究也为后续利用分子标记定位主效QTL提供了参考。2.2 株高、穗长、茎节长及部分产量性状相关性分析
2.3 株高、穗长、节间长数量遗传分析
3 讨 论
4 结 论
猜你喜欢
麦类作物学报(2022年7期)2022-08-09农村百事通(2019年17期)2019-10-08农村百事通(2018年16期)2018-09-29现代农业科技(2017年1期)2017-03-06江苏农业科学(2016年8期)2017-02-15江苏农业科学(2016年8期)2017-02-15中国科技纵横(2016年15期)2016-12-29甘肃教育(2016年22期)2016-12-20江苏农业科学(2016年4期)2016-06-14课程教育研究·下(2016年3期)2016-04-19
