
地理因素对箭竹复合体遗传多样性与遗传分化的影响
2021-06-22 01:20邢晓成张雨曲
西北植物学报 2021年5期
黄 蕾,邢晓成,张雨曲,任 毅
(陕西师范大学 生命科学学院,药用植物资源与天然药物化学教育部重点实验室, 西安 710119)
生物多样性包括遗传多样性、物种多样性和生态系统多样性三个层次,是指全部陆地或水域中生物所拥有的基因、细胞、物种乃至多个生态系统的总称。其中,遗传多样性是生物多样性研究的核心和基础,广义是指地球上所有的生物所携带的全部遗传信息的总和;狭义的遗传多样性则是指种内不同居群间以及居群内的遗传变异,即种内基因的变化[1-3]。一般认为,影响物种水平上的遗传多样性因素可能包括该物种的进化历史、分布范围、繁育系统等,通常认为分布地域广、基因流较强、种子较多的物种其遗传多样性较高,一个物种包含的等位基因越丰富,说明它对环境适应性就越强,进化潜力也越大[4]。
影响植物居群遗传多样性的内部因素包括物种的繁育系统(生殖方式), 遗传漂变, 自然选择, 基因突变[5]和基因流[6], 同时还包括由于环境变化和人为干扰引起的种群隔离和生境片断化[7]等外部因素。内部因素可直接作用于基因组, 引起等位基因数目与频率的变化, 外部因素不会直接改变基因 (等位基因) 的数目与频率, 一般通过间接方式使植物居群的遗传多样性水平和遗传结构发生变化[8-9]。近年来,随着分子标记技术的蓬勃发展,探讨不同环境因子对植物遗传多样性的影响成为领域热点。环境因子一般分为生物因子和非生物因子[10],非生物因子中温度、降水、纬度、海拔等地理因素成为研究者关注的重点[11]。值得关注的是不同植物类群中,同一地理因素与遗传多样性可能呈现出截然相反的关联性。徐振朋等[12]利用15对ISSR分子标记对石竹科裸果木属12个天然居群的遗传多样性进行分析,发现海拔和纬度与遗传多样性呈显著的正相关。苏前[13]基于100个RAPD分子标记对夹竹桃科罗布麻属6个天然居群的研究发现遗传多样性与纬度呈显著正相关,而与海拔呈显著的负相关。因此,为了更好地理解和阐释“地理因素如何影响植物居群的遗传多样性”这一重要问题,需来自更多植物类群的研究实例。
箭竹属隶属于禾本科竹亚科 (Poaceae,Bambusoideae) 北美箭竹族 (tribe arundinariinae) 或木本竹类温带分支, 是竹亚科中的大属之一, 包括80~100种。该属广泛分布于中国西南至喜马拉雅东部的山地, 是大熊猫等珍稀动物的主要食物, 在当地生态系统中发挥重要作用[14]。箭竹属的分类地位和属的范围一直存在争议, 易同培[15-16]曾经依据营养器官性状将箭竹属分为2组6系。然而,与繁育器官相比, 营养器官容易受环境影响而发生变化, 同时营养器官普遍存在趋同演化或平行演化现象。基于已有研究发现,箭竹属内有15个近缘种——箭竹(FargesiaspathaceaFranch.)、矮箭竹(FargesiademissaYi)、毛龙头竹(FargesiadecurvataJ. L. Lu)、拐棍竹(FargesiarobustaYi)、青川箭竹(FargesiarufaYi)、糙花箭竹(FargesiascabridaYi)、缺苞箭竹(FargesiadenudataYi)、华西箭竹[Fargesianitida(Mitford) Keng f. ex Yi)、秦岭箭竹(FargesiaqinlingensisYi et J.X. Shao)、团竹(FargesiaobliquaYi)、伏牛山箭竹(F.funiushanensisYi)、神农箭竹[Fargesiamurielae(Gamble) Yi]、窝竹(Faregsiabrevissima)、细枝箭竹(Fargesiastenoclada)和甜箭竹(Fargesiaostrina)[17],以上物种之间的形态学差异不显著,部分性状存在连续变异,且在已有的系统发育树上位于同一分支[18-19],而箭竹是其中最早命名的物种[20],因此本研究将其称为箭竹复合体(Fargesiaspathaceacomplex)。本研究的前期工作筛选了箭竹复合体的39个天然居群共750个个体,利用14个SSR(微卫星 Simple Sequence Repeat)分子标记,详细探讨了该复合体内的遗传变异式样,并认为其中3个种是成立的,而有4个种显然不成立。同时结合3种居群遗传结构分析方法,将箭竹物种复合体的39个居群分为2个支系(A支系、B支系),其中A支系内部可再划分出华西箭竹亚支系和团竹亚支系,B支系内部可再划分出拐棍竹亚支系、窝竹亚支系、箭竹亚支系和糙花箭竹亚支系,即全部样本分为6个亚支系。另一方面,还发现该复合体内的整体遗传多样性水平并不低,改变了“竹子以营养繁殖为主,遗传多样性水平较低”的传统观念[21]。但是需要指出的是,该研究集中在梳理箭竹复合体内部的遗传结构和居群变异式样,并未详细探讨环境因素,尤其是海拔和纬度对整个复合体内不同支系、亚支系的遗传多样性水平的影响。基于箭竹复合体现有的750个个体大量的遗传变异信息,以及该类群对大熊猫保护领域的重要意义,本研究认为该复合体是深入探讨“地理因素如何影响植物居群的遗传多样性”这一问题的理想材料。
另一方面,遗传多样性可为人们估算植物居群的遗传分化提供重要的基础数据。遗传多样性的计算是基于遗传变异信息,这些遗传变异在居群之间的不均衡分布表现为遗传分化,而遗传分化同样受到地理因素的重要影响[8-9]。因此,本研究在深入分析前期研究工作的箭竹复合体SSR标记数据时,同时关注地理距离与箭竹复合体内的遗传分化水平是否表现出显著的相关性。根据前期野外观察,箭竹复合体内同一支系内的不同类群存在海拔差异,这些类群可能发生了对高海拔区域的适应性分化,目前并不清楚地理距离是否影响了复合体内的适应性分化。综上所述,本研究探讨箭竹复合体的遗传多样性及其与地理因素的关系,旨在揭示箭竹复合体遗传多样性对地理变化的响应,为后续深入研究竹类植物对高山环境的适应性分化奠定基础,并为箭竹复合体内遗传多样性保护和大熊猫主食竹相关研究提供科学依据和理论支持。
1 材料和方法
1.1 微卫星(SSR)引物筛选
本研究的测序样品选择箭竹物种复合体的4个竹种:缺苞箭竹(甘肃省文县)、毛龙头竹(陕西省太白县)、细枝箭竹(四川省彭州县)和伏牛山箭竹(河南省栾川县),用其新鲜叶片,干冰保存送至上海天昊遗传分析中心,利用 Illumina HiSeq 2500平台进行配对末端(paire-end)测序。根据返回的高通量测序信息,在设定文库片段大小时,在主峰基础上±50 bp设定其最大与最小片段范围,其中毛龙头竹(310~410 bp)、缺苞箭竹(280~380 bp)、细枝箭竹(280~380 bp)和伏牛山箭竹(280~380 bp),以双末端方式导入CLC Genomics Workbench 7.5(QIAGEN, Germantown, MD, Germany) 软件中,先去除质量较低的序列,再用从头组装的策略对上一步得到的结果进行组装,所得结果以*.fasta文件格式输出。本研究所筛选14对SSR引物的详细信息,参照黄蕾等文章中相应表格[18]。
1.2 箭竹复合体样本采集
参考《中国植物志》[14]和《中国竹类图志》[22],整理箭竹物种复合体中15个竹种的详细地点,并历时3年对其进行样品采集。采集地点包括四川、陕西、贵州、湖北、河南、甘肃、宁夏等省份。样品采集按照每个居群20~30份DNA材料(由于箭竹物种复合体竹种地下茎属于合轴丛生,地上同一丛不同秆株来自同一个体,因此在样品采集时,选择个体间间隔15 m以上),总共采集39个居群,包括箭竹(2个居群)、矮箭竹(3个居群)、毛龙头竹(7个居群)、拐棍竹(4个居群)、青川箭竹(2个居群)、糙花箭竹(2个居群)、缺苞箭竹(3个居群)、华西箭竹(3个居群)、秦岭箭竹(4个居群)、团竹(1个居群)、伏牛山箭竹(1个居群)、神农箭竹(4个居群)、窝竹(1个居群)、细枝箭竹(1个居群)和甜箭竹(1个居群)。每个居群挑选15~20个个体,共计750个个体进行SSR片段的扩增和测序,详细信息请见黄蕾等文章中相应表格[21]。
1.3 总DNA提取与PCR扩增
利用改良的CTAB[23-24]法提取竹类植物的总DNA, 步骤为: 1)取1.5 g的干燥叶片手工研磨至粉末状,再将粉末状颗粒转入2 mL离心管中,加入800 μL DNA缓冲液Ⅰ,0 ℃保存30 min; 2)离心管放入4 ℃恒温离心机,4 000 r/min恒温离心10 min,弃上清; 3)将离心管取出,加入800 μL已预热的DNA缓冲液Ⅱ(65 ℃),并在65 ℃水浴锅中水浴1 h,间隔15 min轻微摇晃使颗粒与液体充分接触; 4)取出离心管放置于4 ℃恒温离心机中12 000 r/min离心10 min; 吸取上清液至2 mL离心管中,加800 μL的氯仿/异戊醇(体积比24∶1)与80 μL DNA缓冲液Ⅱ,颠倒混合10 min; 5) 4 ℃恒温离心机中常温10 000 r/min离心10 min,取上清液至2 mL离心管中,加入700 μL的氯仿/异戊醇(24∶1)与70 μL DNA缓冲液Ⅱ,颠倒混合10 min; 6)常温10 000 r/min离心10 min,取上清液,加入等体积无水乙醇在-20 ℃冰箱放置1 h;10 000 r/min离心12 min,加入400 μL ddH2O,完全溶解; 7)加入500 μL冰乙醇在-20 ℃放置1 h,12 000 r/min离心10 min,弃上清液,用70%乙醇洗涤沉淀,干燥过夜; 8)加入200 μL ddH2O溶解DNA,-20 ℃保存备用。
PCR扩增首轮反应——10 μL的反应体系,包括酶 2×Es Taq MasterMix 5 μL,ddH2O 4 μL,正反向引物各 0.25 μL,DNA模板0.5 μL。反应程序采用梯度 PCR(TD-PCR)程序:94 ℃ 4 min; 94 ℃ 30 s,62~0.5 ℃, 20 s,72 ℃ 30 s,10个循环; 94 ℃ 30 s, 57 ℃ 20 s,23~24 循环, 72 ℃ 30 s; 72 ℃ 10 min; 25 ℃。第二轮反应——10 μL反应体系,包括酶 2×Es Taq MasterMix 10 μL,ddH2O 8 μL,荧光引物正反向各0.5 μL,DNA模板(首轮反应产物稀释100倍)1 μL。反应程序: 94 ℃ 4 min; 94 ℃ 30 s, 62~0.5 ℃30 s, 72 ℃ 30 s,10个循环; 94 ℃ 30 s, 57 ℃ 30 s, 72 ℃ 30 s, 23~24 循环; 72 ℃ 10 min, 25 ℃。两轮反应中,不同的引物采用不同的荧光进行标记。扩增产物用1.5%琼脂糖凝胶电泳检测后送至江苏金唯智生物科技有限公司进行毛细管电泳检测,使用ABI3730XL测序仪完成荧光检测,得到扩增产物的峰型图谱。
1.4 数据处理与分析
SSR原始数据进行汇总整理后,输入CONVERT 1.3[25]软件进行格式转换,用于后续软件分析。使用POPGENE 1.32软件[26]进行遗传多样性——平均等位基因数(Na)、有效等位基因数(Ne)、香农指数(I)、扩增多态性位点比率(PPL)、期望杂合度(He)、观测杂合度(Ho)的计算。本研究所用的线性回归分析采用R软件包进行计算并绘图[27]。在遗传分化指标上,利用GenAlEx 6.502软件进行Mantel检验[28],通过1 000次迭代运行计算地理距离与遗传距离的相关性。本研究采用的地理距离是基于居群经纬度所得的欧式距离。
2 结果与分析
2.1 海拔和纬度对箭竹复合体遗传多样性的影响
基于750个个体和14对SSR引物的遗传结构分析,39个居群可分为A, B两个支系,A支系可进一步分为2个亚支系,B支系可分为4个亚支系[21]。首先对A, B两个支系的遗传多样性参数——平均等位基因数(Na)、有效等位基因数(Ne)、香农指数(I)、扩增多态性位点比率(PPL)、期望杂合度(He)、观测杂合度(Ho)进行计算,A支系遗传多样性参数的结果为Na=2.93,Ne=1.85,I=0.64,Ho= 0.47,He=0.37,PPL=0.81,B支系为Na=3.05,Ne=1.91,I=0.66,Ho=0.45,He=0.37,PPL=0.86。由图1可见,B支系的遗传多样性略高于A支系。
对6个亚支系的遗传多样性参数进行计算(图2),华西箭竹亚支系内13个居群Na=6.21,Ne=2.05,I=0.84,Ho=0.50,He=0.44,PPL=0.819;团竹亚支系内4个居群Na=4.14,Ne=1.69,I=0.61,Ho=0.37,He=0.33,PPL=0.732;拐棍竹亚支系内4个居群Na=4.57,Ne=1.84,I=0.73,Ho=0.40,He=0.38,PPL=0.857;窝竹亚支系内9个居群Na=5.43,Ne=2.09,I=0.80,Ho=0.43,He=0.41,PPL=0.821;箭竹亚支系内4个居群Na=4.50,Ne=2.08,I=0.76,Ho=0.47,He=0.39,PPL=0.875;糙花箭竹亚支系内5个居群Na=5.50,Ne=2.38,I=1.00,Ho=0.51,He=0.50,PPL=0.929。6个亚支系遗传多样性由高到低的顺序为:糙花箭竹亚支系>华西箭竹亚支系>窝竹亚支系>箭竹亚支系>拐棍竹亚支系>团竹亚支系。在遗传多样性指标上,扩增多态性位点比率PPL在不同亚支系之间变化最小;平均等位基因数Na的变化最大。

图1 箭竹复合体A、B两支系内遗传多样性参数对比Fig.1 The genetic diversity of A and B clades in Fargesia spathacea complex

图2 箭竹复合体6个亚支系组内遗传多样性参数对比图Fig.2 The genetic diversity of six sub-clades in F. spathacea complex
为检测箭竹物种复合体A支系和B支系的遗传多样性与海拔、纬度是否具有显著相关性,将两组各自的遗传多样性指标He与海拔和纬度分别进行线性回归分析,结果如图3所示。A支系的遗传多样性与纬度、海拔均具有显著相关(图3, A支系:R2=0. 5635,r=0.751,P<0.01;R2=0.0151,r=0.123,P<0.01),B支系也可得出同样结论(图3, B支系:R2=0.0700,r=0.264,P<0.01;R2=0.1302,r=0.360,P<0.01)。这表明随着纬度和海拔的增加A支系和B支系的遗传多样性也随之增大。
为检测箭竹物种复合体6个亚支系的遗传多样性与海拔、纬度是否具有显著相关性,将每个亚支系的遗传多样性指标He与海拔和纬度分别进行线性回归分析。拐棍竹亚支系He与纬度的相关性R2=0.3772,r=0.615,P<0.01;He与海拔的相关性R2=0.1948,r=-0.442,P<0.01。窝竹亚支系He与纬度的相关性R2=0.0002,r=0.014,P<0.01;He与海拔的相关性R2=0.6353,r=0.797,P<0.01。箭竹亚支系He与纬度的相关性R2=0.7542,r=-0.868,P<0.01;He与海拔的相关性R2=0.3406,r=0.584,P<0.01。糙花箭竹亚支系He与纬度的相关性R2=0.2314,r=0.481,P<0.01;He与海拔的相关性R2=9E-06,r=0.003,P<0.01。华西箭竹亚支系He与纬度的相关性R2=0.0935,r=0.306,P<0.01;He与海拔的相关性R2=0.1945,r=-0.441,P<0.01。团竹亚支系He与纬度的相关性R2=0.6121,r=0.782,P<0.01;He与海拔的相关性R2=0.0442,r=0.210,P=0.02。由此可见,尽管6个亚支系的遗传多样性和纬度、海拔呈现出显著相关性,但表现出不同的趋势,部分支系表现为负相关。
2.2 地理距离对箭竹复合体遗传分化式样的影响
利用GenAlEx 6.41在箭竹物种复合体水平和A、B支系水平分别进行Mantel检验(图4)。在箭竹复合体水平上,全部39个居群的遗传距离和地理距离未表现出显著的线性关系(R2=0.0032,r=0.057,P=0.11)A支系的检测结果表明遗传距离和地理距离存在显著相关性(R2=0.0613,r=0.25,P=0.01),B支系的检测结果表明遗传距离和地理距离不存在显著相关性(R2=0.0036,r=0.06,P=0.21)。

图3 箭竹复合体A、B两支系遗传多样性水平与纬度、海拔的回归分析图Fig.3 The regression analysis of genetic diversity and latitude, altitude within A and B clade in F. spathacea complex
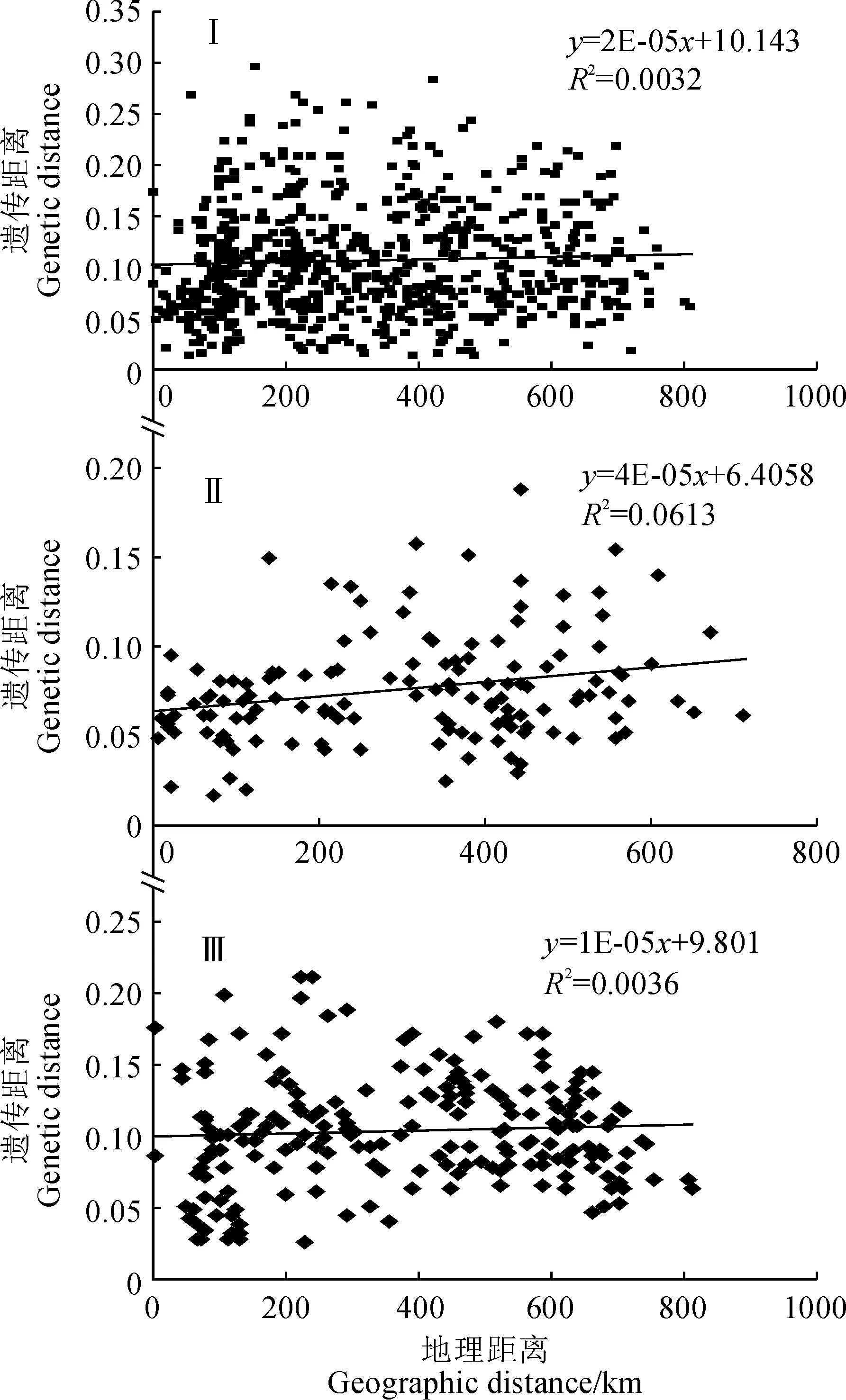
Ⅰ. 箭竹复合体; Ⅱ. A支系; Ⅲ. B支系图4 不同划分组下遗传距离和地理距离 之间的Mantel 检验Ⅰ. Fargesia spathacea complex; Ⅱ. A clade; Ⅲ. B cladeFig.4 The Mantel test of genetic distance and geographic distance under different groups
在6个亚支系水平上, Mantel 检验的结果如图5所示。 拐棍竹亚支系(a)检测结果不显著,相关系数R2=0.3944,r=0.63,P=0.15;糙花箭竹亚支系(b)检测结果不显著,相关系数R2=0.1153,r=0.34,P=0.15;窝竹亚支系(c)检测结果不显著,相关系数R2=0.0558,r=0.24,P=0.10;箭竹亚支系(d)检测结果不显著,相关系数R2=0.0223,r=0.15,P=0.58;华西箭竹亚支系(e)检测结果不显著,相关系数R2=0.011,r=0.11,P=0.40;团竹亚支系(f)检测结果显著,相关系数R2=0.3571,r=0.60,P=0.02。由此可见,B支系内4个亚支系均未表现出遗传距离和地理距离之间的显著相关性,这一结果与前述结果一致;A支系内2个亚支系仅团竹亚支系的遗传距离和地理距离存在较弱的显著相关性。
3 讨 论
影响植物居群遗传多样性的因素包括物种的生殖方式、遗传漂变、自然选择、突变率、基因流,以及种群隔离和生境片断化。另一方面,样本大小和分子标记数量也能对遗传多样性的估算产生一定影响[29-30]。在本研究中,华西箭竹亚支系的居群数目最多,但其遗传多样性并非最高。由此可见,在亚支系水平,样本大小对估算箭竹复合体遗传多样性的影响较小。这表明,箭竹复合体的不同类群在适应不同生境时,遗传多样性水平可能会受到其他生态因子或进化因素的影响。其次,在地理因素与遗传多样性的相关性上,不同物种的研究结果并不一致[9-10]。在本研究中,地理因素与遗传多样性的相关性在不同进化尺度上并不相同。在A、B支系水平上检测出显著相关性,可能是掩盖亚支系之间的遗传结构的结果,并不能反映真实情形。因此建议,开展类似研究工作时应充分考虑遗传结构的影响,先厘清样本内部的遗传分化,再开展多因素与遗传多样性的相关性检测。
在地理距离与遗传距离的相关性上,A支系内包含华西箭竹亚支系和团竹亚支系,本研究结果表明地理隔离在一定程度上影响了其现有分布格局;而B支系内的遗传结构更为复杂,其分布格局可能受其他因素比如自然选择、基因流、遗传漂变等影响。据此推测,地理距离并非是影响箭竹复合体内遗传分化的主导因素。依据本研究前期的发现,B支系下的糙花箭竹和窝竹亚支系内的部分个体检测到明显的杂交迹象,这也为B支系Mantel检验结果不显著提供了合理解释[21]。后续需加强基因流和杂交事件的检测,以明确不同亚支系之间的基因流是否参与了现有遗传分化格局的建立。
本研究通过对箭竹复合体内39个居群的14对SSR数据进行深入分析,发现糙花箭竹亚支系具有最高的遗传多样性,而团竹亚支系的遗传多样性最低。尽管在A、B两个支系水平,遗传多样性和纬度、海拔呈现了显著正相关,但在更精细的亚支系水平,遗传多样性的变化趋势呈现出更为复杂的局面,部分支系表现为负相关。由此推测,箭竹复合体的不同类群在适应不同生境时,遗传多样性可能会受到其他生态因子或进化因素的影响。另一方面,通过Mantel检验结果发现地理距离并非是影响箭竹复合体内遗传分化的主导因素。B支系内的4个亚支系,均未表现出遗传距离和地理距离之间的显著相关性;A支系中仅团竹亚支系的遗传距离和地理距离存在较弱的显著相关性,这表明自然选择、基因流、遗传漂变等因素可能影响了该复合体内的遗传分化。已有结果表明该复合体内部分居群之间可能存在基因流,后续需结合基因流检测推断杂交事件对其遗传分化的影响。鉴于箭竹复合体内不同类群之间存在海拔分化,本研究的结果可为后续深入研究竹类植物的适应性分化奠定基础,亦可为大熊猫主食竹的相关研究提供重要的参考信息。

Ⅰ. 拐棍竹亚支系; Ⅱ.糙花箭竹亚支系; Ⅲ.窝竹亚支系; Ⅳ.箭竹亚支系; Ⅴ.华西箭竹亚支系; Ⅵ.团竹亚支系图5 6个亚支系下遗传距离和地理距离之间的Mantel 检验Ⅰ. F. robusta sub-clade; Ⅱ. F. scabrida sub-clade; Ⅲ. F. brevissima sub-clade; Ⅳ. F. spathacea sub-clade; Ⅴ. F. nitida sub-clade; Ⅵ. F. obliqua sub-cladeFig.5 The Mantel test of genetic distance and geographic distance within six sub-clades
作者贡献:邢晓成和张雨曲采集了全部样本;邢晓成完成了SSR数据的测序和数据分析;任毅教授设计了研究方案;邢晓成和黄蕾完成文章撰写。
猜你喜欢
河北科技师范学院学报(2022年2期)2022-08-26
浙江中医药大学学报(2021年6期)2021-07-12
中国粮油学报(2020年12期)2021-01-09
——以云南墨江自治县为例
贵州民族研究(2019年9期)2019-10-24
神州民俗(2018年9期)2018-11-21
草地学报(2018年5期)2018-11-07
齐鲁周刊(2017年29期)2017-08-08
流行色(2017年1期)2017-05-31
新西部·中旬刊(2016年5期)2016-06-08
测绘学报(2014年2期)2014-01-11
