
缘叶舌蜂与其天敌窄头褶翅蜂的生物学特性及其与环境变量的相关性
2021-07-07 03:52郭鹏飞王明强陈婧婷郭士琨陈国华朱朝东
昆虫学报 2021年5期
郭鹏飞, 王明强, 李 逸, 陈婧婷, 郭士琨,陈国华,朱朝东,4,*
(1.云南农业大学植物保护学院, 昆明 650201; 2.中国科学院动物研究所, 动物进化与系统学(院)重点实验室, 北京 100101;3.中国科学院大学生命科学学院, 北京 100049; 4.中国科学院动物研究所, 农业虫害鼠害综合治理国家重点实验室, 北京 100101)
膜翅目昆虫在生态系统中发挥重要的传粉、捕食和寄生功能。蜜蜂为世界上66%的农作物授粉,直接或间接地对约15%~30%的粮食生产起至关重要的作用(Kremenetal., 2002)。蜜蜂也是野生植物的关键传粉者,即使物种数量的微小变化也会对传粉效率产生重要影响(Garibaldietal., 2013)。隶属于蜜蜂总科(Apoidea)的叶舌蜂是一类典型的传粉昆虫。
叶舌蜂属Hylaeus隶属于膜翅目(Hymenoptera)蜜蜂总科(Apoidea)分舌蜂科(Colletidae),世界已知大约500种,是该科唯一全球分布的属(Kayaalpetal., 2013)。关于这个属的已有研究大多集中于以分类为主的系统学(Magnacca, 2007; Kayaalpetal., 2013)。然而,相关的生物学和生态学,尤其是它们的筑巢习性和天敌种类的研究还很缺乏。
褶翅蜂属Gasteruption隶属于膜翅目(Hymenoptera)褶翅蜂科(Gasteruptiidae),全世界分布500余种,中国已知分布有28种(Zhaoetal., 2012; Chenetal., 2014)。该属物种主要营盗寄生,幼虫以寄主和寄主后代的蜂粮(花蜜和花粉)为食(Boguschetal., 2018)。然而,关于这个属详细的生活史研究还很缺乏。
我们采用了标准化的人工巢管(巢管法)来监测并捕获部分独栖性的膜翅目昆虫。人工巢管作为一个模型系统,已被广泛用于研究独栖筑巢蜂类的生活史(Costa and Gonçalves, 2019)以及保护这些蜂类的多样性(Gastonetal., 2005; von Königslöwetal., 2019)。同时,该方法也可以用于量化分析独栖性蜂所提供的生态服务功能和不同营养级之间的物种互作(Doradoetal., 2011; Staabetal., 2016)。巢管法也可用于果树授粉,如通过巢管诱集壁蜂Osmia等筑巢蜂类为果树和作物授粉,提高坐果率(Vicens and Bosch, 2000; Oliveira and Schlindwein, 2009)。
总之,在基础研究和作物授粉方面,巢管法是一种可信且可量化的方法。膜翅目昆虫通常拥有较短的觅食距离,这使它们与其寄生性天敌对栖息地的环境变化很敏感。因此,人工巢管非常适合研究不同环境梯度变化对膜翅目独栖性昆虫与其寄生性天敌的影响。本研究以亚热带地区江西省分布的缘叶舌蜂Hylaeusperforata及其寄生性天敌窄头褶翅蜂Gasteruptioncorniculigerum为对象,调查并分析了其生物学特性及与生境的关系,以期为更广泛和深入地开展亚热带地区功能性昆虫(传粉者和寄生者)的生物学与生态学研究奠定基础。
1 材料与方法
1.1 研究地点和样地设置
研究地点在江西省德兴市新岗山镇附近的生物多样性与生态系统服务功能实验样地(BEF-China)(29°07′N, 117°54′E, 海拔范围为126~232 m)。实验样地位于亚热带地区,年平均温度16.7℃,年平均降雨量1 821 mm,属典型的亚热带季风气候。样地设计参考Bruelheide等(2014),以25.8×25.8≈667 m2为单位的小样方。每个样方设置了不同的乔木树种丰富度,分别为1, 2, 4, 8, 16和24种。每个样方种植400棵树,彼此水平距离为1.29 m。样方内每个树种的不同个体随机种植在样方内,每种树的个体数相等。我们依据不同的乔木树种丰富度选取了88个小样方进行采样。树种丰富度为1的样方有24个;树种丰富度为2的样方有16个;树种丰富度为4的样方有16个;树种丰富度为8的样方有16个;树种丰富度为16的样方有12个;树种丰富度为24的样方有4个。
1.2 采样方法
我们采用了标准化的人工巢管技术,分别在2015年8月至2018年9月收集缘叶舌蜂的巢穴生物学相关数据。巢管材料选自于当地芦苇,并将其切割成长度为20 cm。将这些直径为0.2~2.5 cm不等的芦苇随机混合放入巢箱,每个样方放置2个巢箱,每月检视样方并把已筑巢的巢管取走,并放入新的巢管。
1.3 室内饲养、生物学数据观测及分类鉴定
将从样方取回的巢管在室内解剖,将每个巢管的样方编号,记录采集日期、巢室长度、巢口直径以及缘叶舌蜂与窄头褶翅蜂的巢室数。并将记录过的巢管放置在试管中,用棉花堵住瓶口,放在室温下饲养;等至巢管内缘叶舌蜂和窄头褶翅蜂成虫羽化。将成虫干制成标本并编号,记录成虫羽化的时间和性别。
根据《浙江蜂类志》和期刊文献Zhao等(2012),使用Olympus双目立体解剖镜将缘叶舌蜂和窄头褶翅蜂鉴定到形态种。证据标本现保存在中国科学院动物研究所进化与系统学(院)重点实验室。
1.4 数据分析
观测原始数据在Excel中记录,在R3.4.3中进行相关性分析。在进行分析前,首先检查了所有环境变量:乔木树种丰富度(tree species richness)、海拔(elevation)、坡度(slope)、北向指数(northness)和东向指数(eastness)是否具有较强的相关性。其中,海拔、坡度、北向指数和东向指数是基于不同样方的数据。同时,为了提高方差齐性,我们将乔木树种丰富度进行了对数转换,并且对所有的环境变量进行了标准化处理(mean=0,SD=1)。缘叶舌蜂和窄头褶翅蜂的多度以及寄生率作为响应变量,使用斯皮尔曼等级相关系数,即Spearman相关系数来检测环境变量与响应变量之间的相关性。
2 结果
2.1 缘叶舌蜂与窄头褶翅蜂生物学特性
共在5个样方的人工巢管中发现缘叶舌蜂40个巢,150个巢室(表1),亲代雌性平均每次建造巢室3.75±2.24个。使用树脂封闭巢口和建造巢室,巢的长度15~175 mm,平均长度74.46±39.16 mm;巢的直径3~6 mm,平均直径4.73±1.31 mm。后代巢室位置按照雌性在内雄性在外分布,并且雌性羽化时间晚于雄性2~3 d。后代雌性数量远高于雄性,雌雄性比2.57;雌性成虫体长7.1~7.3 mm,雄性成虫体长6.8~7.1 mm。缘叶舌蜂的亲代雌性在每年5-9月筑巢;一年多代;后代成虫羽化时间在每年4-8月;8月和9月筑巢的缘叶舌蜂当年不会羽化,以末龄幼虫越冬,羽化时间为翌年的4月底;后代食用的花蜜有柠檬香味。
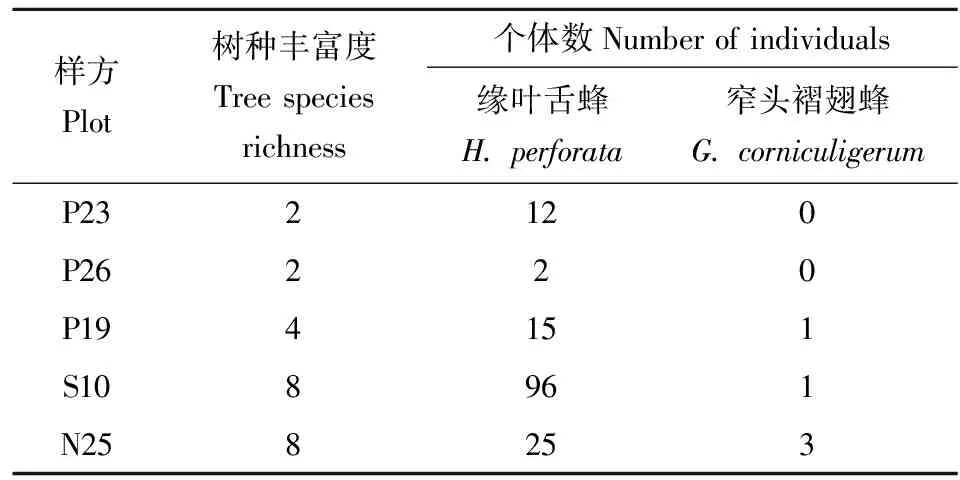
表1 缘叶舌蜂与窄头褶翅蜂采样数据(江西德兴, 2015年8月至2018年9月)
共发现5头窄头褶翅蜂(表1),包括4头雌性和1头雄性。雌性成虫体长16.1~16.8 mm,雄性成虫体长15.9 mm。窄头褶翅蜂营盗寄生;只发现此蜂在7月寄生缘叶舌蜂,9月羽化活动;发育时间超过其寄主1个月;羽化时间远晚于同期筑巢的缘叶舌蜂(7月筑巢的缘叶舌蜂8月羽化,而7月寄生的窄头褶翅蜂全部在9月羽化)。
2.2 环境变量对缘叶舌蜂与窄头褶翅蜂的影响
缘叶舌蜂的多度与乔木树种丰富度呈显著正相关[Spearman相关系数(RS)=0.95,P<0.05](图1: A),而其他环境变量对缘叶舌蜂的多度没有显著影响(表2)。窄头褶翅蜂仅在较高乔木多样性的样方中出现(表1)。窄头褶翅蜂的多度与乔木树种丰富度呈显著正相关(RS=0.92,P<0.05)(图1: B),而其他环境变量对窄头褶翅蜂的多度没有显著影响(表2)。各环境变量对窄头褶翅蜂的寄生率没有显著影响(表3)。

图1 树种丰富度与缘叶舌蜂(A)和窄头褶翅蜂(B)多度的关系

表2 缘叶舌蜂和窄头褶翅蜂的多度与环境变量的相关性

表3 窄头褶翅蜂的寄生率与环境变量的相关性
3 讨论
3.1 缘叶舌蜂生物学特性
本样地巢管收集到的其他蜜蜂类群如丘切叶蜂Megachilemonticola、拟丘切叶蜂M.pseudomonticola、双叶切叶蜂M.dinura、粗切叶蜂M.sculpturalis、壮壁蜂Osmiataurus和脊跗拟孔蜂Hoplitiscarinotarsa均为一年1代。它们在当年筑巢后,只会在次年羽化。而缘叶舌蜂的筑巢活动时间较为广泛。叶舌蜂属的其他种类如H.alcyoneus不发生滞育,以成虫越冬(Paini and Roberts, 2005)。而我们实验观察到的缘叶舌蜂是以末龄幼虫越冬。与巢管内其他筑巢膜翅目比较,该物种后代偏向于更多的雌性。经典的费希尔理论认为后代性比应大致相等(Paini and Bailey, 2002),而缘叶舌蜂性比偏向的原因目前尚不清楚。同时,我们也收集到了此种蜂储备的花粉,有待进一步分析,以明确其蜜源植物。
3.2 窄头褶翅蜂生物学特性
截止目前,未见关于窄头褶翅蜂的生物学报道。之前关于此物种的研究仅限于分类学,并未报道相关的生物学(Zhaoetal., 2012)。本样地巢管收集到的其他盗寄生蜂类群如Chrysisprincipalis、Chrysiskoma、Chrysidinaesp.、Coelioxysfenestrata、厚腹尖腹蜂Coelioxyscrassiveutris和基翅腹蜂Euaspisbasalis均在寄主的单个巢室中产下一粒卵。它们的后代会在寄主的巢室内完成生长发育,并不破坏其他巢室,并且羽化时间与寄主接近。而窄头褶翅蜂的幼虫会将缘叶舌蜂巢内的所有巢室破坏,吃完所有寄主后代的蜂粮,并且羽化时间远晚于寄主。原因可能是窄头褶翅蜂相比于其他盗寄生类群,消耗了更多的能量,而且体型也较大,所以生长发育时间更长。但是本研究并未观察到缘叶舌蜂的后代是被此种的亲代还是后代幼虫杀死。同时,本研究也未观察到窄头褶翅蜂详细的寄生行为。对褶翅蜂属的其他种类也有一些生物学行为研究(Cruz Macedoetal., 2012; Boguschetal., 2018),但是这些研究并未报道类似的寄生现象。
3.3 环境对缘叶舌蜂和窄头褶翅蜂的影响
植物物种丰富度与节肢动物类群之间存在着普遍的正相关关系(Bassetetal., 2012; Schuldtetal., 2019)。本研究也发现缘叶舌蜂和窄头褶翅蜂的多度与树种丰富度呈正相关(图1)。寄主的可利用性是所有寄生性天敌的主要资源,寄生性天敌的多度和寄生率通常更多地取决于寄主多度,而不是环境因素(Ebelingetal., 2011; Staabetal., 2016)。虽然有时也会出现广食性寄生蜂,如Melittobiasp.,但大多数寄生蜂都是寄主特异性的,只攻击特定地区的一个或几个密切相关的物种(Tylianakisetal., 2007)。本研究发现窄头褶翅蜂只寄生缘叶舌蜂。东向坡在一天的早晨就能得到直射的阳光,从而迅速变暖,有助于独栖筑巢膜翅目昆虫的觅食和后代发育(Staabetal., 2016)。但是,本研究并未发现缘叶舌蜂和窄头褶翅蜂与东向指数的相关性(表2和3)。仅在5个样方收集到缘叶舌蜂,我们猜测缘叶舌蜂的食物与筑巢材料可能来源于这些样方中特定的植物。由于本研究收集的一些物种,主要是寄生类群还未被鉴定,所以关于环境变量对膜翅目类群的整体影响还有待于进一步分析。
3.4 结论
本研究着重探讨了中国亚热带地区森林生态系统中的缘叶舌蜂与其天敌窄头褶翅蜂的生物学特性,同时分析了环境对缘叶舌蜂与窄头褶翅蜂多度的影响,发现缘叶舌蜂是典型的独栖性蜜蜂,一年多代,在春夏初秋时期活动,后代雌性数量远多于雄性,以末龄幼虫越冬。窄头褶翅蜂的盗寄生方式不同于巢管发现的其他盗寄生类群,通常会消耗掉寄主储存的所有蜂粮,并且羽化时间远长于寄主缘叶舌蜂。本研究发现缘叶舌蜂和窄头褶翅蜂的多度与树种丰富度呈显著正相关。
致谢感谢BEF-China的设计者Martin-Luther-Universität Halle-Wittenberg的Helge Bruelheide教授、University of Zurich的Bernhard Schmid教授和中国科学院植物研究所马克平研究员;感谢景德镇学院杨波博士和薛玉洗先生及中国科学院植物研究所李姗博士和刘晓娟博士对实验站的维护;感谢新岗山镇齐银泉先生在采样中给予的帮助;感谢中国科学院动物研究所袁峰老师和中山大学陈华燕博士对本实验缘叶舌蜂和窄头褶翅蜂的鉴定。
猜你喜欢
资源导刊(信息化测绘)(2022年4期)2022-11-24
乡村科技(2021年17期)2021-10-20
安顺学院学报(2021年4期)2021-09-16
安徽林业科技(2021年2期)2021-06-15
电脑爱好者(2020年10期)2020-07-28
电脑爱好者(2019年16期)2019-10-30
小溪流(画刊)(2017年6期)2017-07-10
中国科技纵横(2016年15期)2016-12-29
青岛科技大学学报(社会科学版)(2015年4期)2016-01-25
商业经济研究(2009年26期)2009-10-27
