
不同光照强度对赤皮青冈幼苗光合特性的影响
2021-07-30 02:15王佩兰李志辉朱中华
中南林业科技大学学报 2021年7期
夏 婵,李 何,王佩兰,李志辉,王 波,朱中华
(1.中南林业科技大学 林学院,湖南 长沙 410004;2.江西省林业科学院,江西 南昌 330032;3.攸县林业局,湖南 攸县 412300)
幼苗阶段是植物生长发育周期及更新过程中的薄弱环节[1]。研究发现,光照是影响幼苗存活、生长,幼树建植及森林更新的重要环境因子[2]。近年来,有关光环境影响林下幼苗更新及特定植物群落繁育的理论日益得到关注[3]。众多研究表明,当光照条件受限时,植物会发生一系列形态结构响应及生理生态响应,尤其是光合生理响应以提高自身竞争能力和生存适合度。研究发现,凤丹白幼苗在夏季高光条件下,遮阴可有效缓解光抑制现象,发生积极的光合响应[4];而油桐幼苗在遮阴条件下净光合速率显著降低,发生消极的光合响应[5]。因此,当光照条件受限时,不同幼苗光合生理响应机制在存在显著差异,对不同光照强度下幼苗光合特性进行探索,适度人工干预林下幼苗光环境的选择,有助于保持天然植物群落的更新及延续。
赤皮青冈Cyclobalanopsis gilva为壳斗科Fagaceae 青冈属Cyclobalanopsis的常绿阔叶乔木,该树种材质硬度高且纹理漂亮,是一种很有发展前景的高档用材树种;同时,赤皮青冈树形高大雄伟且为深根性树种,在涵养水源、保持水土方面也有较好表现,是兼具园林观赏、生态绿化与工业用材等多功能于一体的珍贵乡土树种[6]。赤皮青冈在我国南方多省均有分布,由于人类活动的干扰,天然起源的赤皮青冈优势群落已十分稀少,现存赤皮青冈多以古木大树散落分布于其他群落。目前关于赤皮青冈的研究主要集中在地理分布、引种栽培、数量分类[7]、干旱胁迫下赤皮青冈的光合生理响应机制、赤皮青冈育苗造林技术[8]等方面,关于不同光照强度下赤皮青冈幼苗的响应研究仅局限于生长指标,对不同光照强度下赤皮青冈幼苗光合特性的研究仍未见报道。因此,本研究以1年生赤皮青冈幼苗为试验材料,通过不同密度遮阳网实现5 种光照强度,探究赤皮青冈幼苗光合色素含量、光合参数及叶绿素荧光参数在不同光照强度下的变化趋势,旨在确定赤皮青冈幼苗的最佳生长光照条件,为赤皮青冈幼苗培育及人工干预林下幼苗更新提供理论依据。
1 材料与方法
1.1 试验地概况
试验地位于湖南省东北部汨罗市林业科技示范园(112°51′~113°27′E,28°28′~29°27′N),该地区属于典型的亚热带湿润性气候,年平均气温为17℃;年平均日照时数为1 650.1 h;年平均降水量为1 289.8~1 556.2 mm。
1.2 试验设计
本试验以生长状况良好且长势一致的1年生赤皮青冈幼苗为材,平均地径为4.23 cm,平均苗高为29.12 cm,采用29 cm×25 cm×27 cm 的聚乙烯塑料盆对赤皮青冈幼苗进行单株定植。盆栽基质选用苗圃表层黄心土,并施入定量的基肥。
根据试验地附近赤皮青冈生境光照强度的变化范围,试验设置5 个处理:全光照为对照(0遮阴)、普通黑色2 针半遮阳网(30%遮阴)、加密黑色4 针遮阳网(50%遮阴)、加密黑色6针遮阳网(70%遮阴)、普通黑色10 针遮阳网为处理(90%遮阴),分别记为CK、T1、T2、T3、T4。其中,遮阴率于晴朗天气下用便携式光量子测定仪进行确定,全光照条件下光照强度为1 296 μmol·m-2s-1,各遮阴棚光照强度分别为904、654、397、132 μmol·m-2s-1。遮阴棚架构采用林场间伐所得杉木进行搭建,并用铁丝进行加固处理,最后用对应针数遮阳网对遮阴棚进行全覆盖,四周垂下部分距离地面40 cm 高度不进行遮阴,以确保遮阴棚内外空气流动自由。苗期正常管理,确保不同处理间栽培管理技术一致。试验指标测定均在遮阴处理后40 d 进行。
1.3 试验指标测定方法
1.3.1 光合色素含量的测定
采用浸提法[9]进行测定。随机取各遮阴处理下健康无损的赤皮青冈幼苗叶片,用蒸馏水洗净叶片表面污物,去中脉,将叶片剪成0.1~0.2 mm 的细丝,称取0.1 g 于试管中,加入按95%乙醇和无水丙酮1∶1 标准配置的浸提液10 mL,置于黑暗环境下浸提24 h,期间每隔4~6 h 轻晃样品以保证充分浸提。用紫外分光光度计对浸提液进行测定,分别记录其在波长663、645 和470 nm 下的吸光值,根据公式计算光合色素含量的值。
1.3.2 光响应曲线的测定
于2020年6月中旬晴朗无风天气9:00—11:00,使用Li-6400 便携式光合仪进行光响应曲线测定。选择红蓝LED 光源,控制参比室CO2浓度为400 μmol·s-1,流速500 μmol·s-1,光强梯度设置为:2 000、1 800、1 500、1 200、900、600、300、200、150、100、75、50、25、0 μmol·m-2s-1。每个处理选择3 株长势良好的赤皮青冈幼苗,对其同一生长部位的成熟叶片进行测定,重复3 次,光响应特征参数选用叶子飘的非直角双曲线模型[10]进行拟合计算。
1.3.3 光合气体交换参数的测定
于2020年6月中旬晴朗天气9:00—11:00,使用Li-6400 便携式光合仪进行气体交换参数进行测定。选择透明叶室,设置CO2浓度为400 μmol·s-1,流速为500 μmol·s-1,光强为1 000 μmol·m-2s-1。每个处理选择3 株长势良好的赤皮青冈幼苗,对其同一生长部位的成熟叶片进行测定,重复3 次。
1.3.4 荧光参数的测定
于2020年6月中旬晴朗天气6:00—9:00,使用MINI-PAM 便携式荧光仪进行叶绿素荧光参数测定。测定前用叶夹对待测叶片进行暗处理,时长控制在30~40 min,每个处理选择3 株长势良好的赤皮青冈幼苗,对其同一生长部位的成熟叶片进行测定,重复3 次。
1.4 数据分析
采用Excel 2016进行数据整理和绘图,SPSS 19.0软件进行单因素方差分析及相关性分析。
2 结果与分析
2.1 不同光照强度对赤皮青冈幼苗光合色素含量的影响
由图1可知,赤皮青冈幼苗叶片叶绿素a(Chl a)、叶绿素b(Chl b)、总叶绿素(Chl t)、类胡萝卜素(Car)含量随遮阴程度的加深而增加,均在T4 处理下达到最大,分别比CK 高103.43%、174.60%、122.27%、101.69%,且T2、T3 和T4 处理的光合色素含量均与CK 差异显著(P<0.05),Chl a/b 则随遮阴程度的加深呈现出先升后降的趋势,在T4 处理下最小,仅为CK的75.72%。
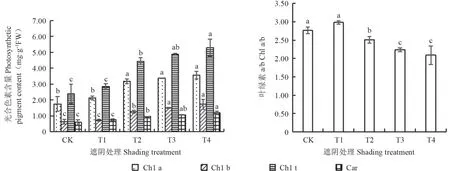
图1 不同光照强度下赤皮青冈幼苗叶片光合色素含量Fig.1 Photosynthetic pigment content of Cyclobalanopsis gilva seedling leaves under different light intensity
2.2 不同光照强度对赤皮青冈幼苗光响应曲线的影响
由图2可知,各组之间响应曲线都有较大差异,同一光合有效辐射(PAR)下,Pn由大到小分别为T2>T1>CK>T3>T4;当0<PAR<200 μmol·m-2s-1时,Pn增速较快,各处理之间差异较小;在200~600 μmol·m-2s-1阶段,Pn增速变缓;在1 200 μmol·m-2s-1以后,CK、T1、T2 处理下赤皮青冈幼苗净光合速率逐渐趋于稳定,而在T3 和T4 处理有下降的趋势,表明T3、T4 处理下赤皮青冈幼苗可能已经产生光抑制。
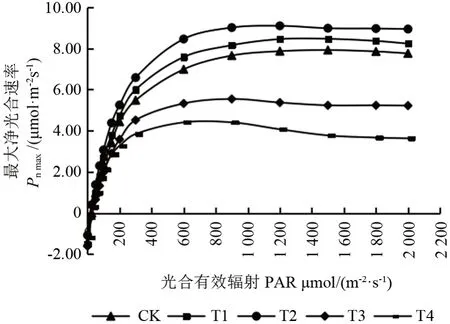
图2 不同光照强度下赤皮青冈幼苗叶片光响应曲线Fig.2 Light-response curves of Cyclobalanopsis gilva seedling leaves under different light intensity
由表1可知,不同遮阴处理对赤皮青冈幼苗最大净光合速率(Pnmax)、光饱和点(PLS)、光补偿点(PLC)、暗呼吸速率(Rd)均有显著影响(P<0.05)。Pnmax随遮阴程度的加深而呈现出先升后降的趋势,表现为T2>T1>CK>T3>T4,且各处理组之间差异显著(P<0.05),T2 最高,为CK 的116.69%,T4 最低,仅为CK的55.13%;随遮阴程度的加深,PLS逐渐降低,且各处理组之间差异显著(P<0.05),CK 分别为T1、T2、T3、T4 的106.78%、116.90%、144.51%、198.39%;PLC与Rd均在T2 处理下达到最小值,分别为18.86、1.06 μmol·m-2·s-1,仅为CK 的79.10%、69.75%;各处理组间表观量子效率(AQY)变化不明显。

表1 不同光照强度下赤皮青冈幼苗叶片的光响应特性†Table 1 Light response characteristics of Cyclobalanopsis gilva seedling leaves under different light intensity
2.3 不同光照强度对赤皮青冈幼苗光合气体交换特性的影响
由图3可知,遮阴对赤皮青冈幼苗各气体交换参数均有显著影响。随着遮阴程度的加深,净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)、均呈现出先升后降的趋势,Pn、Gs均表现为T2、T3、T4 处理与CK 处理差异显著(P<0.05),且均在T2 处理下达到最大,T2 处理分别为CK 处理的115.91%、137.50%;Ci仅在T2处理与CK 差异显著(P<0.05),T2 最大,为CK 的107.37%;各处理组蒸腾速率(Tr)与CK均 差异显著(P<0.05),T2 最大,为CK 的126.62%,T4 最小,仅为CK 的45.45%。

图3 不同光照强度下赤皮青冈幼苗叶片的气体交换参数Fig.3 The gas exchange parameters of Cyclobalanopsis gilva seedling leaves under different light intensity
2.4 不同光照强度对赤皮青冈幼苗叶绿素荧光参数的影响
由表2可知,遮阴对赤皮青冈幼苗PS II 的实际光合效率(Y(II))与PS II 的相对电子传递速率(ETR)影响显著(P<0.05),Y(II)与ETR 变化规律一致,均表现为T2>T1>T3>CK>T4,T2 处理Y(II)和ETR 显著高于CK(P<0.05),分别为CK 的123.81%、124.69%;遮阴对光化学猝灭系数(qP)影响显著(P<0.05),T1、T2、T4 处理qP 均与CK 差异显著(P<0.05),T2 最高,为CK 的114.29%,T4 最低,仅为CK的85.71%;CK 处理PS II 调节性能量耗散的量子产量(Y(NPQ))与其余各组之间差异不显著;各处理组间PS II 的最大光合效率(Fv/Fm)的值均处于0.80~0.82。
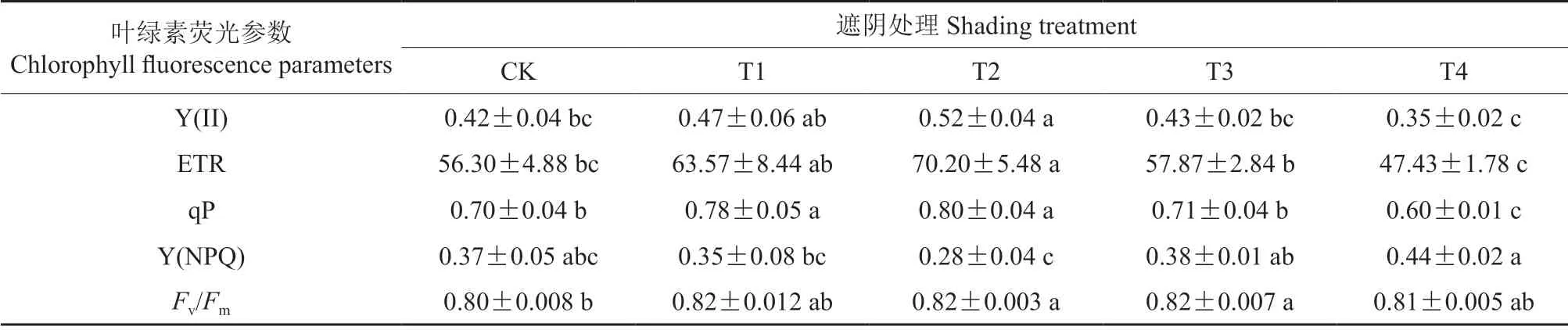
表2 不同光照强度下赤皮青冈幼苗叶片的叶绿素荧光参数Table 2 Chlorophyll fluorescence parameters of Cyclobalanopsis gilva seedling leaves under different light intensity
2.5 不同光照强度下赤皮青冈幼苗光合生理指标相关性分析
由表3可知,Pn与Chl a/b、Pnmax、PLS、Gs、Ci、Tr、Y(II)、ETR、qP 呈显著正相关关系,与Chl t、Y(NPQ)呈显著负相关关系。其中PLC与Rd、Y(II)与ETR 均呈显著正相关关系。
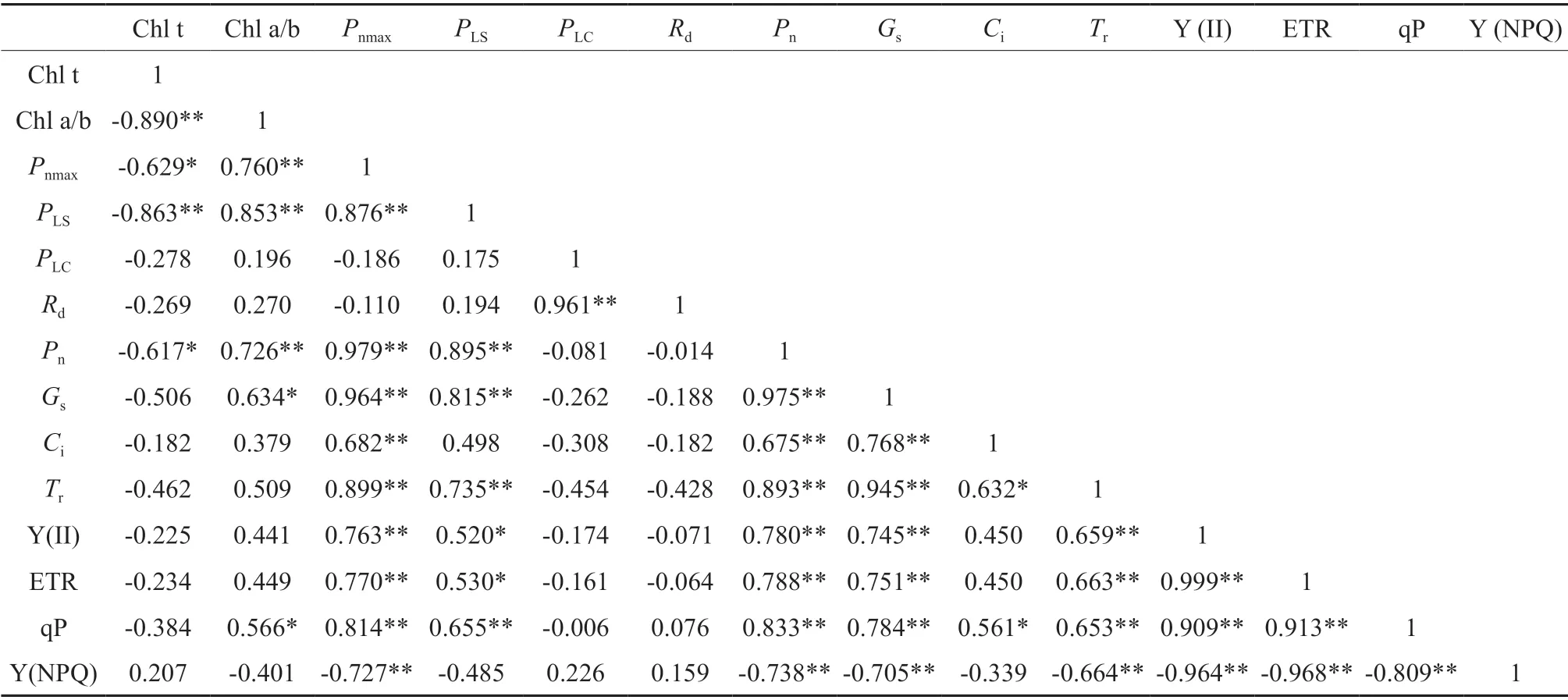
表3 不同光照强度下赤皮青冈幼苗光合生理指标的相关性分析†Table 3 Correlation analysis of photosynthetic physiological indexes of Cyclobalanopsis gilva under different light intensity
3 结论与讨论
3.1 结 论
综上所述,赤皮青冈幼苗通过调节光合色素含量、光合参数及叶绿素荧光参数以适应变化的光环境,表现出一定的光合可塑性。研究结果表明,不同光照条件下赤皮青冈幼苗的光合特性存在显著差异。与全光照相比,适度遮阴(30%~50%遮阴)条件下赤皮青冈幼苗光合能力得到改善,且在50%遮阴条件下光合强度达到最高,而重度遮阴(70%~90%遮阴)则导致赤皮青冈幼苗光合能力受到限制。因而,今后在进行赤皮青冈幼苗培育及人工干预林下幼苗更新时,应注意光照环境的选择,50%遮阴为赤皮青冈幼苗的最适生长光照条件。本文对不同光照强度下赤皮青冈幼苗光合特性的研究仅局限于控制环境条件测量,今后研究中应加强非控制环境条件的测量,以全面阐述赤皮青冈幼苗在不同光照强度下的光合响应机制。
3.2 讨 论
3.2.1 不同光照强度对赤皮青冈幼苗光合色素的影响
叶绿素、类胡萝卜素在植物光合作用进程中执行光能的吸收、传递及转化等功能[11]。光合色素含量及Chl a 与Chl b 的比值一定程度上反映了植物是否适应所处的光照环境[12]。本研究中,赤皮青冈幼苗光合色素含量随遮阴程度的加深而递增,与CK(全光照)处理相比,Chl a、Chl b、Chl t 和Car 在T2(50%遮阴)、T3(70%遮阴)和T4(90%遮阴)处理下均显著增加,赤皮青冈幼苗光合色素含量在弱光环境下表现出积极的适应特征。叶绿素a 主要吸收红光,叶绿素b 主要吸收蓝紫光,遮阴条件下红光比例降低,蓝紫光比例升高[13],Chl a/b 降低是植物利用弱光能力的判别标准[14]。本研究中,赤皮青冈幼苗Chl a/b 随遮阴程度的加深而先升后降,在T4 处理下达到最低,表明在遮阴条件下赤皮青冈幼苗弱光利用能力的机制得到明显改善。
3.2.2 不同光照强度对赤皮青冈幼苗光合参数的影响
通过拟合光响应曲线以得到一系列光响应特征参数,对于了解植物光合作用进程中的光化学效率有着重要意义[15]。本研究中,各处理下赤皮青冈幼苗光响应特征明显不同。PLS和PLC代表植物对光照强度耐受的上限和下限,分别反映植物对强光和弱光的利用能力[16]。在各遮阴处理下赤皮青冈幼苗PLS均显著低于CK,在遮阴条件下更早达到PLS,说明赤皮青冈幼苗在遮阴条件下对光的利用效率得到有效提升。Rd暗呼吸速率是指植物维持自身生命活动所必需消耗体内有机物的效率[17]。在PAR 相同的情况下,T1(30%遮阴)和T2 处理具有较高的Pn,说明赤皮青冈幼苗在适度遮阴条件下发生积极的光合适应。其中,T2 处理与CK 相比,PLC及Rd显著降低且PLC、Rd与Pn呈负相关关系(表3),表明此时赤皮青冈幼苗通过降低PLC以提高对弱光的利用效率,同时通过降低Rd以保证植物体内营养物质的有效积累,表现出较强的光合生理调节能力,有利于在有限的光照条件下最大程度利用低光量子密度,满足植物的生长需求。Pn是体现植物光合效率最直接的指标,T3 与T4 处理下Pn显著下降,说明过低的光照条件下赤皮青冈幼苗光合作用机制不能正常运转,产生弱光胁迫,但T3 与T4 处理下光合色素含量显著增加,且Pn与Chl t 呈显著负相关关系,说明光合速率是一系列光化学反应综合作用的结果,光合色素含量并非植物光合作用能力的唯一限制因素[18],其含量高低不能直接代表植物净光合速率。
具有一定光合可塑性的植物可以通过调节其气体交换参数来适应变化的光环境。气孔是植物与外界进行水分和CO2等气体交换的主要通道,直接影响到植物光合作用进程[19]。本研究中,赤皮青冈幼苗Pn、Gs、Ci随遮阴程度的加深均表现出先升后降的趋势,Pn与Gs、Ci、Tr呈显著正相关关系(表3),在T2 处理下,各气体交换参数均显著高于CK 处理,表明适度遮阴条件下赤皮青冈幼苗通过开放气孔来维持较高的Tr从而有效提高Pn以满足植物自身生长需要。与CK 相比,T3与T4 处理下Gs、Ci、Tr均显著下降,说明重度遮阴条件下赤皮青冈幼苗CO2和水分进出气孔阻力变大,此时气孔限制因素是Pn下降的主要原因。
3.2.3 不同光照强度对赤皮青冈幼苗叶绿素荧光参数的影响
叶绿素荧光分析技术被称为快速、准确测定植物光合生理状况的理想探针,包含丰富的光合作用信息[20]。Y(II)表示PS II 反应中心在部分关闭情况下的实际原初光能捕获效率。ETR 表示PS II 反应中心在实际光强条件下的表观电子传递效率,其值是根据Y(II)值与PAR 计算得出:ETR=PAR×Y(II)×0.84×0.5[21]。本研究中,Y(II)与ETR 呈显著正相关关系(表3),不同处理下均表现为T2>T1>T3>CK>T4,重度遮阴时Y(II)与ETR 显著降低,说明在过低光照条件下赤皮青冈幼苗光能捕获效率与电子传递能力下降,光合作用受限。qP 是PS II 反应中心天线色素吸收的光能用于光化学电子传递的份额,该值的大小可以反映PS II 电子传递活性的高低[22]。qP 在T1和T2 处理下显著高于CK,且Pn与qP 呈显著正相关关系(表3),说明此时PS II 反应中心电子传递活性显著提升,光合电子传递处于最佳状态。Y(NPQ)是光保护的重要指标,反映植物将PS II天线色素吸收的过剩光能耗散为热能以保护光合机构的能力[23]。Y(NPQ)在T2 处理下达到最小,说明在适度遮阴条件下赤皮青冈幼苗显著降低了光合色素所吸收的光能用于保护性耗散比例,将更多的能耗集中分配于光化学反应。因此,赤皮青冈幼苗在适度遮阴条件下通过提高PS II 反应中心光化学活性并降低热耗散的方式以提高光能转化效率,保证苗木的正常生长发育。
猜你喜欢
中国动物保健(2022年10期)2022-11-04
少儿科技(2022年2期)2022-03-05
数学大王·趣味逻辑(2021年6期)2021-09-27
烟台大学学报(自然科学与工程版)(2021年1期)2021-03-19
阅读(科学探秘)(2020年8期)2020-11-06
小资CHIC!ELEGANCE(2019年40期)2019-12-10
绿色科技(2019年2期)2019-05-21
女性天地(2016年10期)2017-04-25
水禽世界(2015年6期)2016-03-04
水禽世界(2014年4期)2014-09-21
