
模拟氮沉降对喀斯特原生乔木林土壤氮矿化的影响
2021-07-30 02:15赵文君刘永涛谭成江崔迎春侯贻菊刘延惠丁访军
中南林业科技大学学报 2021年7期
赵文君,刘永涛,谭成江,崔迎春,吴 鹏,侯贻菊,刘延惠,丁访军
(1.贵州省林业科学研究院,贵州 贵阳 550005;2.中国电建集团贵阳勘测设计研究院有限公司,贵州 贵阳 550081;3.茂兰国家级自然保护区管理局,贵州 荔波 558400)
由于矿物燃料燃烧,大量工业氮肥使用等使大气中含氮化合物激增,大气氮沉降也不断增加(从1.5 kg·hm-2a-1增加到4.2 kg·hm-2a-1)[1],氮沉降作为驱动因子势必改变森林土壤生态过程,直接影响了森林土壤氮素转换方向、速度和通量[2],森林土壤氮矿化、硝化和反硝化作用作为氮素循环的重要环节,与森林生态系统氮可利用性、各形态比例、养分利用效率相关联[3-4],与森林群落演替、植物多样性、生态系统退化和健康等之间存在反馈关系,是决定森林生态系统功能和结构的关键过程[5],因此研究氮沉降下森林土壤氮矿化的动态变化对了解氮素周转、营养循环及森林生态系统生产力等具有重要意义[6]。
喀斯特生态系统有特殊的水文地质二元结构及复杂的峰林、峰丛、漏斗和洼地等地貌形态,小尺度范围内形成石面、石沟和土面等小生境,不同小生境异质性显著,进而引起小气候、土壤及其他生态因子的变化[7-8],可能对土壤氮库动态及净矿化速率产生影响。目前在非喀斯特区常态地貌不同生态系统及植被类型下开展的相关研究结论并不一致,如包翔等[9]研究表明氮沉降显著增加了大兴安岭白桦次生林土壤氨化速率、硝化速率和矿化速率,其中氨化速率增幅大于硝化速率。方运霆等[10]研究表明南亚热带森林土壤有效氮随氮沉降的增加而增加,且随施氮时间增长,施氮样方和对照样方差距越大。而闫聪微等[11]研究发现高氮处理显著降低了中亚热带森林土壤NO3--N、NH4+-N 含量。Chappell 等[12]研究发现多年氮添加并没有引起沿海花旗松林土壤氮矿化速率的增加。且有关喀斯特森林小生境尺度下氮沉降对土壤氮矿化的影响的研究基本没有涉及,因此,本研究选择茂兰喀斯特原生林为研究对象,对模拟氮沉降下土面和石沟小生境土壤无机氮库及净氮矿化速率动态变化进行比较,探讨喀斯特森林小生境下土壤氮矿化特征及对氮沉降的响应,旨在为进一步研究全球气候变化对森林生态系统氮循环的影响提供基础资料。
1 材料与方法
1.1 研究区概况
试验地点位于荔波县茂兰国家级自然保护区,地理坐标107°52′12″~108°05′42″E,25°09′27″~25°20′40″N,属于世界上同纬度地区留存下来仅有的、原生性强、相对稳定的喀斯特森林生态系统,是喀斯特区原生性森林常绿落叶阔叶混交林分布面积最大的地区[13],其自然状况参见文献[14-16]。该区顶极森林群落原生乔木林,乔—灌—草结构完整,乔木为主,林分密度1 022 株/hm2,林分高度10~20 m,覆盖率80%以上,主要优势种有青冈栎Cyclobalanopsis glauca、圆果化香Platycarya longipes、卵果海桐Pittosporum lenticellatum、硬斗石栎Lithocarpus hancei、丝栗栲Castanopsis fargesii等[16]。试验区属于典型的喀斯特地貌,土壤以石灰土为主,土层浅薄(20 cm左右)且不连片,多存在于岩石裂隙或出露岩石形成的沟地中,土壤的理化性质、剖面形态等区别于地带性土壤[13,17]。
1.2 研究方法
1.2.1 样地设置与样品采集
通过对贵州荔波喀斯特森林生态系统定位观测研究站原有固定样地踏查,筛选了有代表性的原生乔木林群落样地,海拔770 m,坡度20°~25°,中下坡位,样地面积30 m×30 m。按照刘方等[17]对小生境划分,考虑小生境面积和数量,样地内随机选取了石沟(出露的岩石溶蚀沟或侵蚀沟)、土面(面积相对较大的小型台地)2 种小生境3 m×3 m 布置模拟氮沉降试验。0~15 cm 土壤理化性质,土面的有机质、全N、全P 和全K 含量分别为102.30、4.57、0.91、和16.63 g/kg,pH 值为6.20;石沟的分别为108.02、4.95、1.02 和17.11 g/kg,pH 值为6.56。施氮量设置3 个处理,分别为低氮N1(25 kg·hm-2a-1)、高氮N2(50 kg·hm-2a-1)和对照N0(0 kg·hm-2a-1)。土面、石沟每个处理均为3 个重复。每个样方间距>3 m,以防止相互干扰。自2018年6月开始施氮试验,施氮频率为每月1 次,施氮时间为1 a,按照处理水平的要求,将各样方所需NH4NO3溶解至1 L 水中,用喷雾器在该水平样方中均匀喷洒,每次施氮量相当于全年模拟总施氮量的1/12,对照样方喷施等量的水,以减少因外加水而造成的试验差异。本研究采用原状土原位培养连续取样法来估测净氮矿化速率,每月施氮前采样,先去除地表凋落物,将2 支长15 cm、内径7.5 cm 的PVC 管打入土壤,一支土柱低温保存带回实验室,测定NH4+-N、NO3--N 含量的初始值;另一支管顶用通气但不透水的塑料薄膜封闭,管底用纱布封口后进行原位培养,培养30 d 后取出PVC 管,同时将下一批管按上述方法布置于前一次培养点附近,每个处理样方内均重复取样3 次。
土壤铵态氮、硝态氮的测定分别采用氧化镁—扩散法[18]和酚二磺酸比色法[18]。
1.2.2 数据处理
土壤氮转化率计算公式如下:
M1=[I1-I0]/D
式中:M1为土壤净氨化、净硝化、净氮矿化速率,mg/(kg·d);I0和I1分别为培养前后铵态氮、硝态氮、矿化氮含量,mg/kg;D为培养天数,d。
所有数据用Excel 2010 软件处理及作图,SPSS 16.0 软件统计分析,采用单因素方差分析检验不同小生境和不同氮处理间无机氮含量、土壤矿化速率的差异显著性。
2 结果与分析
2.1 模拟氮沉降对土壤NH4+-N 含量的影响
各小生境土壤NH4+-N 含量存在明显季节变化(图1),添加外源氮后,在3—4月改变了土壤NH4+-N 含量的变化趋势。生长季含量高于非生长季,10月达到全年的峰值,4月降到谷值。从整个试验期看,与对照相比,添加外源氮增加了土壤NH4+-N 含量(3月除外),且随着氮水平的升高而增加,但不同月份各小生境增加程度不一。就土面而言,N2 处理除3月外都显著增加了土壤NH4+-N 含量(P<0.05),其中5月最大增加了67.12%。N1 处理仅9月和5月土壤NH4+-N 含量显著增加(P<0.05),5月最大增加了36.86%,其余月份增加了6.99%~30.58%。石沟生境下,N2 处理除3月和4月外其余月份均达到了显著水平(P<0.05),5月最大增加了67.53%。N1 处理在7、12月和翌年1、5月NH4+-N 含量显著增加(P<0.05),5月最大增加了32.88%,其余月份增加了10.43%~29.02%,效果不显著。NH4+-N含量及对氮添加的响应都是石沟略高于土面。

图1 不同氮沉降水平土面和石沟土壤-N动态Fig.1 -N dynamics in soil surface and stone gullyat different nitrogen deposition levels
2.2 模拟氮沉降对土壤NO3--N 含量的影响
添加外源氮基本上不会改变土壤NO3--N 含量的变化,各生境土壤NO3--N 含量先骤升到高值而后降到低值,随后波动上升,均在9月达到峰值,土面和石沟分别于10月和12月降到低值,5月NO3--N 含量积累达到小高峰。N 添加均会不同程度的增加土壤NO3--N 含量,且是随着氮水平的增加而增加。土面中,N2 处理在7—9月、11月和翌年5月会显著增加土壤NO3--N 含量(P<0.05),其中8月最大增加了99.92%。其余月份与对照相比增加了44.72%~57.77%,但作用效果不显著。N1 处理仅在8、9月使土壤NO3--N 含量显著上升(P<0.05),此外均有不同程度的上升(14.89%~43.84%),但都未达到显著水平。石沟实施氮处理后土壤NO3--N 含量8月才呈现出显著增加的态势(P<0.05),略晚于土面。N2 处理在8—11月、翌年1、5月显著增加土壤NO3--N 含量(P<0.05),其中9月最大增加了72.97%。其余月份与对照相比增加了24.59%~68.78%,没有达到显著水平。N1 处理仅在8—9月使土壤NO3--N 含量显著上升(P<0.05),其余使土壤NO3--N 含量上升(7.28%~24.77%),但作用效果不明显。对照中NO3--N 含量石沟显著高于土面(P<0.05),而氮添加后,NO3--N 增加程度土面大于石沟。
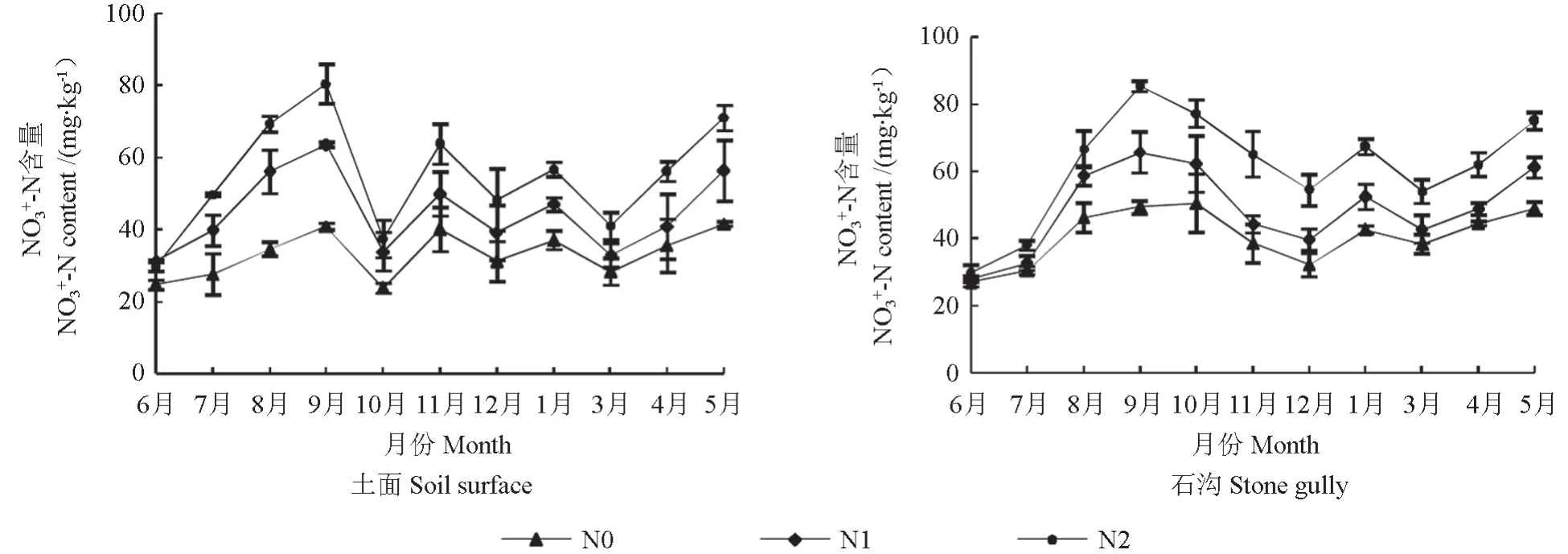
图2 不同氮沉降水平土面和石沟土壤NO3--N 动态Fig.2 NO3--N dynamics in soil surface and in stone gully at different nitrogen deposition levels
2.3 模拟氮沉降对土壤氮氨化速率的影响
各生境土壤净氨化速率存在显著的季节差异(P<0.05),开始阶段6—7月为正值最大,而后出现负值,在11—12月达到最小,正净氨化速率和负净氨化速率交替出现。土面、石沟对照中净氨化速率分别为-1.03~1.09 mg/(kg·d),-1.01~0.97 mg/(kg·d),氮添加后,与对照相比,净氨化速率都呈现增长趋势,但差异不显著。净氨化速率表现为石沟>土面,但两者生境间差异不显著。
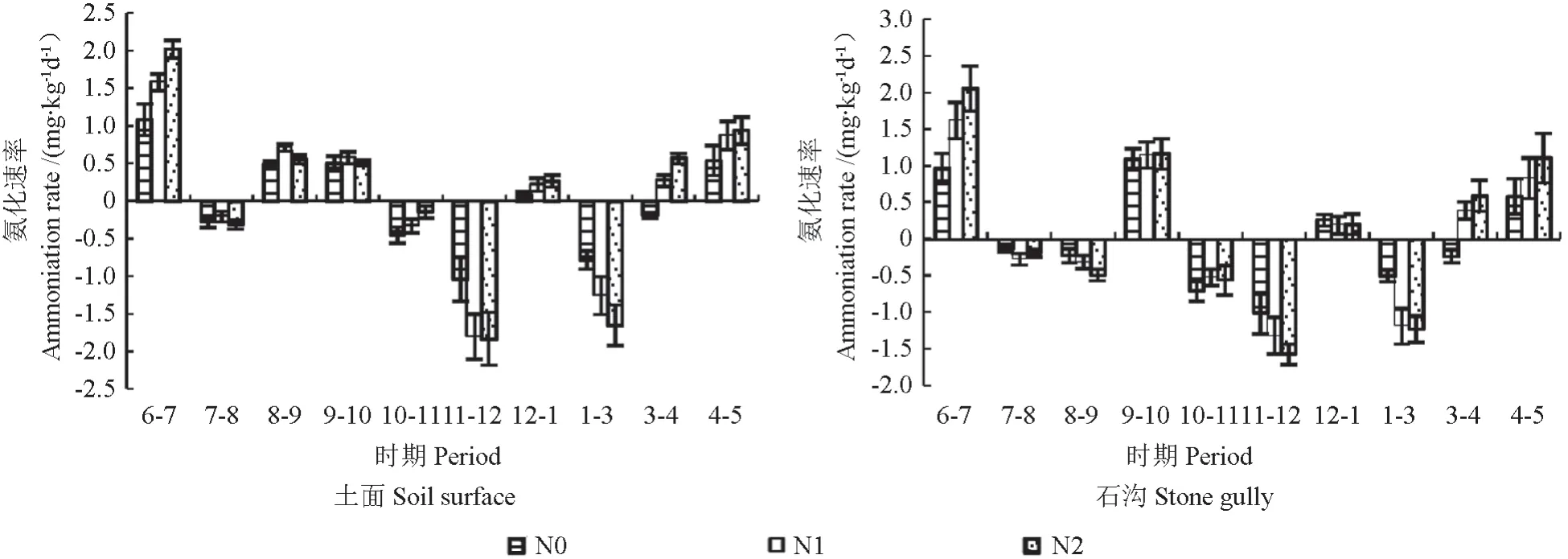
图3 不同氮沉降水平下土面和石沟净氨化速率动态变化Fig.3 Dynamic changesof soil net ammonification rate of on the soil surface and stone gully at different nitrogen deposition levels
2.4 模拟氮沉降对土壤氮硝化速率的影响
各生境净硝化速率存在明显的季节动态(P<0.05),都在7—8月达到最高,土面9—10月达到最低值,石沟最弱发生在10—11月。硝化速率出现负值可能是部分铵态氮被微生物吸收转化为有机氮,或土壤反硝化作用增强所致。整个氮沉降过程中,与对照相比,硝化速率也呈增长趋势。土面和石沟对照中净硝化速率为-0.57~0.54 mg/(kg·d)、-0.40~0.52 mg/(kg·d),由于森林土壤硝化作用来源于氨化作用的NH4+-N,氨化作用直接影响硝化速率的变化,硝化速率往往低于氨化速率[2],氮添加激发效应对NH4+-N 促进作用高于NO3--N。氮添加对净硝化速率的影响并不明显,土面和石沟的生境净硝化速率差异不显著。
2.5 模拟氮沉降对土壤氮矿化速率的影响
各小生境净矿化速率季节间差异显著(P<0.05),净矿化速率在6—7月最高,生长季初中期都为正值,11—12月达到最低为负值,翌年1—3月也是处于低值,表示土壤以固持为主。氮添加后土壤净矿化速率都呈增长趋势,但没有达到显著水平。净矿化速率对氮沉降的响应石沟略大于土面。

图4 不同氮沉降水平下土面和石沟净硝化速率动态变化Fig.4 Dynamic changesof soil net nitrification rate on the soil surface and stone gully at different nitrogendeposition levels
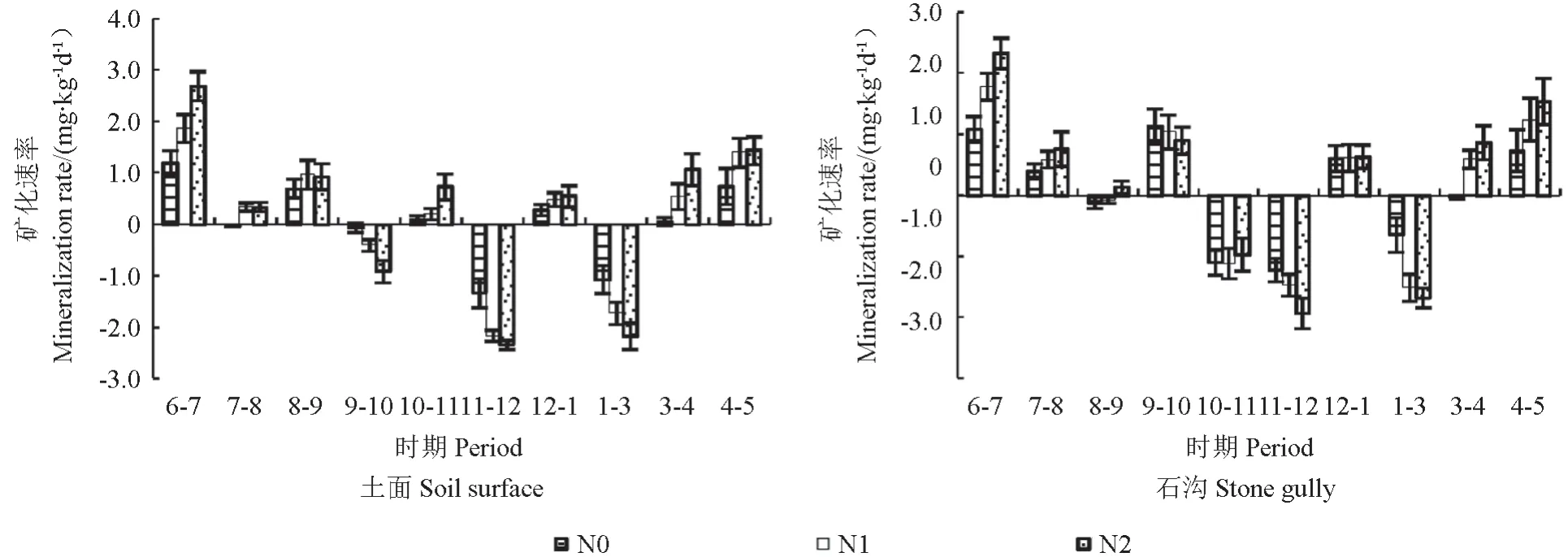
图5 不同氮沉降水平下石沟净氮矿化速率动态变化Fig.5 Dynamic changesof soil net mineralization rateon the soil surface and stone gully at different nitrogen deposition levels
3 结论与讨论
3.1 模拟氮沉降下不同小生境无机氮库的动态变化
整个培养期间,与对照相比,不同小生境不同氮添加均增加了土壤NH4+-N、NO3--N 含量,高氮处理的增幅高于低氮处理,与胡艳玲等[19]、方运霆等[10]、包翔等[9]和向元彬等[20]的研究结果一致。一是因为外源氮提高了微生物活性,氮沉降的激发效应加速了土壤有机物的矿化,土壤C/ N因过量的N 和有机物结合而降低,促进了土壤有机物分解和养分释放[21-22]。二是氮输入增加了土壤和凋落物层的矿质氮含量,使植物吸收与硝化菌、反硝化菌对氮的竞争得到了缓冲,硝化、反硝化作用增加,进而使土壤有效氮增加[22]。此外施加NH4NO3直接增加土壤有效氮水平。
土面和石沟之间有效氮差异不显著,主要是因为各小生境间土壤温度的差异不足以对土壤矿化产生显著影响。其次喀斯特森林土面和石沟有效氮含量本底值较高,供氮能力强[16],显著高于南亚热带林[10,20]、温带白桦林[9]等森林,本研究施氮量及施氮时间未能引起土壤氮矿化的显著变化。土面和石沟土壤有效氮含量均集中在9—10月,土壤有效氮月动态是植物生长与土壤微生物活动共同作用的结果,原因在于两者均受温度和水分控制[19]。在一定阈值内随温度和湿度升高,微生物和酶活性高,凋落物分解快,加速了氮矿化过程,增加土壤有效氮含量。冬季1月有所回升,有研究表明冬季低温土壤仍具有明显的氮矿化过程[4,23],且冬季为非生长季,植物对氮素的需求较少,有效氮大量富集于土壤中[22]。另外冬季降水量少,土壤水分低,带负电荷的硝态氮不易在离子交换中流失,导致冬季硝态氮含量较高[24]。
3.2 模拟氮沉降下不同小生境土壤净氮矿化特征
土面和石沟的小生境土壤净氮矿化速率变化趋势基本一致,表现为生长季初中期高且为正值,生长季末期较低且为负值,与土温的季节变化密切相关。温度对土壤微生物活性的影响非常显著,一定温度范围内土壤的矿化速率一般都与温度正相关[25]。本研究中土温最高的6—7月土壤净矿化速率达到最高值。
土壤氮矿化速率对氮沉降的响应一般而言短期内随着施氮量增加而增加[26],本研究氮添加后土壤净氨化速率、净硝化速率、净矿化速率与对照相比呈现增长趋势,与Magill 研究结果一致[27],美国缅因州东部针阔混交林地土壤的氮素净矿化量进行了9年的原位观测,结果表明低剂量(50 kg·hm-2a-1)和高剂量(150 kg·hm-2a-1)NH4NO3输入均显著促进土壤氮素净矿化通量,并且高剂量氮处理的促进作用更为明显,外源氮输入量对土壤净矿化通量的贡献率约为64%。因为增加的氮被土壤有机质吸收,导致 C/N 减小,从而提高了分解过程中氮的释放速度[28]。或添加的无机氮被微生物固定,从而促使原来有机氮的矿化和释放[29]。
也有一些森林研究表现为高氮处理抑制土壤氮矿化[29],或随施氮时间增长,氮矿化速度在到达高峰后开始降低,降低到接近对照或低氮处理的水平,有的甚至更低[11,30]。Aggangan 等[31]研究表明施入氮加快了氮素各形式的周转速率,使土壤的矿化潜力减小。也有解释是由于土壤微生物产生的腐殖质分解酶的量随土壤有效氮含量增加而减少,从而降低矿化速率[30,32]。此外还与森林生态系统的氮状态是否饱和密切相关[2]。本研究没有出现降低的趋势,是因为喀斯特森林生态系统土壤矿化过程以硝化作用为主导,由于硝态氮易于流失或发生反硝化作用引起气态损失[16],降低了有效氮在土壤中的积累,以至本试验施氮强度和施氮时间对土壤的影响还没达到该生态系统“阈值”。
氮添加对不同小生境影响不同,石沟响应高于土面。不同的小生境中温度、湿度及凋落物供给的差异等最终是通过影响到林下不同的微生物类群的数量、种类和活力来影响氮素矿化[28]。研究发现,外加氮源对土壤氮的激发效应总趋势是肥力高的土壤高于肥力低的土壤[16,28]。石沟土壤有机质、全N、全P、全K 含量要高于土面。土壤净氮矿化速率一般与土壤C/N 成反比[30],石沟土壤C/N 要低于土面,C/N 低有利于有机质分解矿化。但土壤氮矿化过程受土面、石沟小生境影响不显著,可能是因为喀斯特森林微地貌小尺度空间的土壤、小气候等生态因子的差异不足以引起土壤氮矿化的显著变化。
本研究为期1年,模拟氮沉降仅处于初期阶段,喀斯特森林生态系统有效氮未达到饱和阈值,仍存在氮素施肥效应,氮输入增加了土壤硝化作用速度和硝态氮流失的潜力,降低了该生态系统氮留存,说明喀斯特森林生态系统对氮沉降有一定的缓冲能力,利于该生态系统稳定、健康、长远发展。喀斯特生境复杂异质性大,小生境类型多样,本研究只选择了石沟和土面2种小生境类型,且试验时间短,有一定的局限性。今后,可增加施氮水平梯度,适当提高高氮水平,进一步研究喀斯特森林生态系统各小生境长期氮沉降响应与适应机制,同时可加强与常态地貌不同生态系统之间的对比研究,更好地探究森林生态系统氮沉降背景下的运转规律,以期为森林生态系统的可持续发展提供理论基础和管理依据。
猜你喜欢
水土保持学报(2022年5期)2022-10-10
科学技术与工程(2022年22期)2022-09-29
农业机械学报(2022年7期)2022-08-08
耕作与栽培(2022年2期)2022-07-01
建材发展导向(2022年6期)2022-04-18
中学生数理化(高中版.高一使用)(2021年3期)2021-06-09
环境与发展(2019年12期)2019-02-12
中学化学(2016年10期)2017-01-07
科学与财富(2016年7期)2016-03-25
绿色科技(2014年5期)2014-08-08
