
重楼属植物分子生物学研究进展
2021-08-05 06:43杨青淑严靖婷段宝忠
中草药 2021年15期
杨青淑,王 婧,江 媛, 杨 燕,严靖婷,段宝忠*
1.大理大学药学院,云南 大理 671000
2.云南省科学技术院,云南 昆明 650228
重楼属ParisL.为广义百合科植物,是重要的药用类群,有2000 多年的药用历史。全世界重楼属植物有29 种,主要分布于欧亚大陆温带和热带地区。历版《中国药典》收载七叶一枝花P.polyphyllaSmith var.chinensis(Franch.) Hara.及滇重楼P.polyphyllaSmith var.yannanensis(Franch.) Hand.-Mazz.作为重楼药用。重楼具有清热解毒、消肿止痛、凉肝定惊之功效[1],是云南白药、宫血宁胶囊、季德胜蛇药片等著名成药的主要原料。由于重楼药材市场需求巨大,但其药用部位生长周期长,自然繁殖效率低,加之无序采挖和过量应用,致使该属多个物种濒临灭绝,已被列为禁止出口物种,其资源的稀缺,已成为制约相关医药工业可持续发展的瓶颈。近年来,随着分子生物学技术的不断发展,出现了许多新的研究热点,重楼属植物的分子生物学研究是目前的热点之一。国内外学者采用分子生物学技术对重楼属植物开展了相关研究,在遗传多样性、系统分类、功能基因等方面取得了诸多成果,为重楼属植物的鉴别、种群遗传多样性、亲缘关系、植物生长发育和代谢调控机制等相关研究,提供了新的思路和参考。目前已有学者对其遗传多样性进行了研究[2],但有关其功能基因、分子鉴定、系统进化等研究领域尚缺乏系统的总结,限制了对重楼属植物的进一步开发和应用。鉴于此,从分子标记、分子鉴定、转录组学、蛋白质组学、功能基因等方面,对重楼属相关分子生物学研究成果进行了综述,并就当前存在的问题进行探讨,以期为该属植物资源的保护和可持续利用提供科学参考。
1 分子标记
1.1 遗传多样性和亲缘关系分析
遗传多样性研究对揭示重楼属物种的进化、种质资源利用和生物学保护策略的制定具有重要意义。近年来,国内外学者利用简单重复序列(simple sequence repeat,SSR)、分子标记技术(inter-simple sequence repeat,ISSR)、EST-SSR(expressed sequence tag-simple sequence repeats)、扩增的限制性内切酶片段长度多态性( amplified fragment length polymorphism,ALFP)、特异引物PCR 标记随机扩增多态性 DNA(random amplified polymorphic DNA,RAPD)等分子标记方法,对重楼属植物的遗传多样性开展了深入研究[3-10]。基于分子标记的遗传多样性统计发现,重楼属植物多态位点百分率(percentage of polymorphic loci,PPL)达79.1%~100%,表明遗传多样性丰富,尤其是对华重楼、黑籽重楼和滇重楼的研究较为深入。利用SCoT(start codon pargeted polymorphism)对重楼属多个物种的研究表明,不同物种间遗传多样性水平为七叶一枝花>宽叶重楼>具柄重楼>狭叶重楼>宽瓣重楼>北重楼[11]。此外,多位学者基于多种分子标记对重楼的亲缘关系进行了研究,如基于CDDP(conserved dna-derived polymorphism)分子标记的非加权组平均法(unweighted pair group method arithmetic averages,UPGMA)分析表明[12],滇重楼与长药隔重楼之间的遗传距离较远,分别聚到了不同的组,这与ISSR[13-14]和限制性位点扩增多态性(restriction site amplification polymorphism,RSAP)[15]的研究结果一致;而黑籽重楼、无瓣黑籽重楼单独聚为一组,七叶一枝花、狭叶重楼聚为一组的结果,与SCoT 标记的分析结果不一致,即黑籽重楼与七叶一枝花聚为一组,狭叶重楼单独聚为一组[16]。两者结论不同,可能与所用的群体种源以及来源不同有关,其亲缘关系有待进一步研究证,实相关信息见表1。
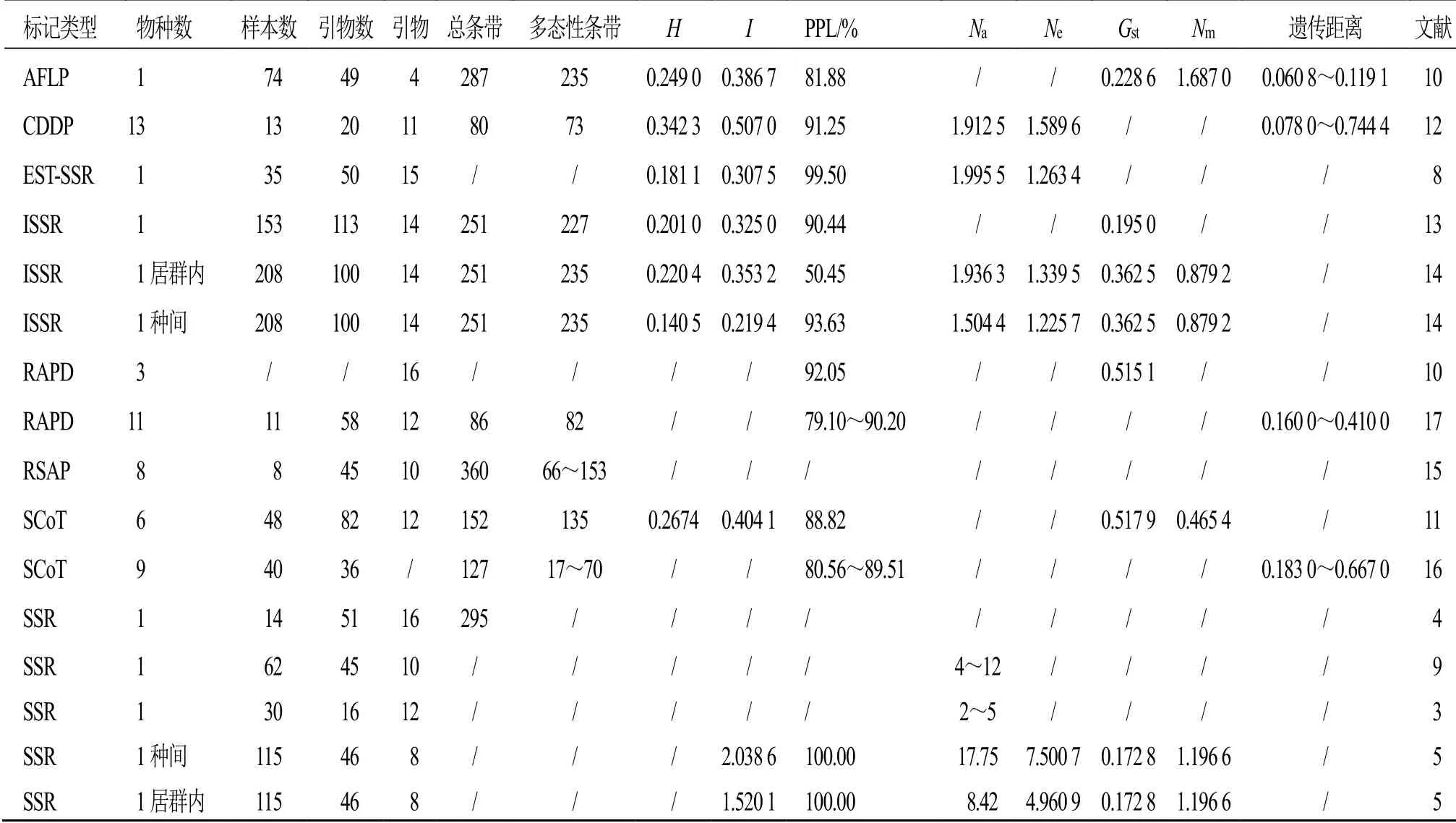
表1 重楼属植物遗传多样性参数Table 1 Genetic diversity parameters of plants from genus Paris
1.2 种质资源和种群研究
优良的种质是重楼属植物资源保护和开发利用的重要保证。在物种间水平的研究上,唐荣华等[17]利用RAPD 对川滇地区的野生重楼11 个种群进行研究,结果12 个引物产生86 个RAPD 标记,其中82 个具有多态性,11 个物种的多态性高于85.2%;李壮等[16]及程虎印等[11]分别对的重楼属植物遗传多样性进行研究,发现四川和陕西的重楼属植物多态性高于80.56%和88.82%,上述研究表明重楼属物种水平上的多样性较高,且与起源地区相关。在物种内种群水平上,研究表明七叶一枝花[18]和黑籽重楼[19]种群内和种群间的遗传多样性变异大体相当;滇重楼和多叶重楼居群内的遗传多样性不足,但居群间具有较高的遗传多样性[13-14],推测生境片段化所带来的遗传漂变,以及地理隔离造成生态地理环境的差异,可能是各居群间出现较高遗传分化的主要原因之一[14]。然而,与之相反,亦有多项研究发现黑籽重楼、多叶重楼的变种滇重楼和七叶一枝花等重楼属植物的种群间,其居群内遗传分化偏高,大于居群间分化[5,10,20],如滇重楼只有30.85%的分子变异存在于居群间,而有69.15%的变异存在于居群内[10];在七叶一枝花中有37.26%的分子变异存在于居群间,62.74%的变异存在于居群内[10];在黑籽重楼中有22.86%的遗传变异存在于居群间,77.14%的遗传变异存在于居群内[20]。这一现象可能由于重楼属植物特殊的繁育方式有关,其能自交结实的特性增加了居群间的分化[20-21],同时人为因素是导致重楼资源匮乏和居群破坏、分布零星的直接原因之一[5]。上述分析表明,重楼属植物种群,特别是滇重楼和华重楼种群仍处于“濒危”生境中,重楼属核心种质资源的建立和保存仍有待加强。
2 分子鉴定
2.1 基于多重聚合酶链反应(PCR)法的分子鉴定
PCR 扩增指纹图谱技术直接用通用引物对基因组进行PCR 扩增,通过扩增条带指纹的差异进行鉴定。它具有不需要已知DNA 序列信息,信息位点多等特征[22]。Duan 等[23]利用高分辨溶解曲线法(high resolution melting,HRM),成功开发了重楼法定基原植物及其混伪品的有效鉴定方法,鉴定准确率为100%。研究者通过分析滇重楼、华重楼与其同属近缘种的ITS序列差异,设计特异性引物,开发了基于PCR技术,准确鉴定《中国药典》法定基原物种及其混伪品[24-25]。上述方法无需进行测序,实现了重楼及其混伪品的有效、快速鉴定。
2.2 DNA 条形码鉴定
近年来,多位学者对重楼属植物的DNA 条形码研究进行了探讨,主要涉及matK、psbA-trnH、ITS2、rpoB、rpoC1、rbcL等序列。刘涛等[26]采用psbA-trnH序列对12 个重楼品种进行分析,发现该序列可区分滇重楼及其同属物种。Ji 等[27]利用ITS、psbA-trnH和trnL-trnF等序列单独或联合对重楼属进行系统发育分析,发现重楼为一个属而不是3 个属的分类(DaiswaRafinesque、KinugasaTatewaki & Suto 和ParisLinnaeus)。姜黎等[28]应用matk、trnL-trnF、rpoC1和ITS序列,鉴别重楼不同近缘品种以及常见伪品,结果表明ITS 序列作为DNA 条形码候选序列,ITS+trnL-trnF 组合序列作为其补充序列,可以有效鉴别重楼药材不同近缘种以及药材混伪品。研究还发现,ITS 与其他序列相比,其在重楼属物种中的鉴定效率更高[29-31]。此外,有学者认为ITS2序列能够准确鉴定重楼属药用植物,可以作为潜在的药用植物通用条形码序列[32-33],基于ITS2 序列构建NJ 树能有效区分重楼属药用植物[34]。方海兰等[35]采用ITS 序列对重楼种子种苗的鉴定进行了研究,发现其可有效鉴别正品重楼及其混伪品的种子种苗。叶方等[36]采用ITS2 序列对重楼属植物进行种内分子鉴定,发现该属植物在NJ(neighborjoining)树中能被明显区分开,并建议将狭叶重楼作为七叶一枝花的变型处理。过立农等[37]采用NJ 树、ITS 和ITS2 2 个序列相结合的方法,发现其可有效将60 批重楼样品鉴别为宽瓣重楼、毛重楼、七叶一枝花、花叶重楼、独龙重楼、五指莲重楼、西藏延龄草、平伐重楼、华重楼和南重楼。该研究为重楼属药材的鉴别开辟了一条新途径。
3 系统进化与谱系地理学
3.1 基于全叶绿体基因组的系统进化
随着分子生物学和基因测序技术的发展,叶绿体基因组越来越受到广大科研人员的青睐。基于叶绿体基因组的丰富数据,分类学家对不同分类阶元的系统发育关系进行了重建,为重楼属植物的系统发育问题提供了新的证据。Do 等[38]首次测序并拼接北重楼的全叶绿体基因组。Huang 等[39]对11 个重楼属物种的叶绿体全基因组进行了测序拼接,并结合北重楼进行分析,发现所研究样本的cemA基因均为假基因,且除四叶重楼和北重楼外,其余重楼的叶绿体基因组的结构十分保守。Gao 等[40]对花叶重楼和禄劝花叶重楼进行重新测序和拼接,并对它们的基因组结构特征进行了描述。杨丽芳[41]对33 种重楼属植物的全叶绿体基因组和核糖体DNA 序列进行测序,发现重楼属植物的叶绿体基因组高度保守,均为一个闭合环状DNA 分子,且rDNA 序列和全叶绿体基因组构建的系统树结果有明显冲突。李晓娟等[42]对华重楼及百合目植物全叶绿体基因组进行了比较,获得华重楼全叶绿体基因环结构,发现编码序列中的终止密码子可以区分华重楼和北重楼,同时华重楼的cemA基因是假基因,推测cemA结构及假基因化现象对重楼属植物的进化与系统发育具有重要意义。
3.2 基于DNA 条形码的系统进化
ITS 是被子植物中应用较广的一种分子标记方法,除用于物种鉴定外,已成功应用于重楼属植物的系统进化研究。翁周[43]对多叶重楼ITS 序列进行比对和聚类分析,发现多叶重楼的ITS 序列比较保守,11个多叶重楼样本明显分为3 支,发现小重楼与其他变种的关系较远,滇重楼和原变种的关系较近,而与其他变种的关系相对较远。唐荣华等[44]比较15 种重楼植物和Trillium tschonoskiiMaxim.的ITS 序列,认为重楼属可划为ParisLinnaeus、KinugasaTatewaki & Suto 和DaiswaRafinesque 3 个独立的属。而唐铭霞[45]对重楼属植物21 个种及变种ITS 区序列以及trnL-F基因间区分析,支持把重楼属归为1 个属ParisLinnaeus。上述研究为重楼属植物的分子鉴定提供了重要的理论基础。
3.3 谱系地理学研究
李恒[46-47]对重楼属的细胞学和地理分布格局进行研究,认为重楼属植物存在2 种基本核型:热带核型和温带核型,其起源于上新世(或更早)的亚洲大陆滇、黔、桂地域。其中,云贵高原至四川邛崃山区为该属的多样化中心。Ji 等[27]根据该类群原始种类(北重楼亚属ParisAubgen.)的分布情况及高等植物的核型演化规律,推测重楼属起源于中新世前期东亚的日本、朝鲜半岛和我国东北、华北附近的地域,由低纬度地区逐渐扩散进入中国的亚热带地区。该研究起源地居群分化为北重楼亚属.的祖先,扩散到中国亚热带地区的居群分化为蚤休亚属EuthyraAubgen.的祖先,因云贵高原和横断山地区复杂多变的地理环境和气候条件,形成该属的多样化中心。Huang 等[39]基于重楼属11 个种构建的系统发育树表明,蚤休属DaiswaRaf.中分布于华东、华中、华南和中南半岛地区与分布于中国西南和喜马拉雅地区的重楼属植物形成了2 个独立的谱系,认为该分布格局的独特性可能与这2 个谱系间经历了长期的隔离分化有关。此外,杨丽芳[41]认为由于环太平洋的造山运动(渐新世/中新世界线附近)、亚洲夏季风加强(晚中新世以来)、青藏高原隆升(晚中新世到早上新世)、冰期/间冰期循环(更新世)等气候和地质变化,创造了高度隔离和多样性的生境,促进了重楼属植物的谱系多样性形成。
4 功能基因组学研究
4.1 转录组学研究
转录组学可揭示基因表达与一些生命现象之间的内在联系,为深入了解重楼属植物发育和进化具有重要意义。多位学者采用高通量测序技术,对滇重楼进行分析,发现了多个参与甾体皂苷生物合成、激素信号、种子休眠等有关的基因[48-50]。南京农业大学张成才等[51]采用转录组测序方法研究华重楼种子的休眠解除机制,发现赤霉素处理是解除华重楼休眠的必要条件。随着对miRNA 在调控植物生长发育中的作用的认识不断深入,杨金龙[52]和Ling 等[49]对滇重楼的种子和种皮中的miRNA 文库进行了测序,首次提供了已知的miRNA 图谱及其靶点,这对进一步研究滇重楼种子休眠的分子机制和重楼属植物相关基因的功能研究提供了重要科学依据。
4.2 蛋白组学研究
蛋白质组学的概念最早由Marc Wilkins 在1994年提出,标志着生命科学进入后基因时代[53]。目前有关重楼属植物的蛋白组学研究,主要集中在基因表达调控成分代谢领域。Liu 等[54-55]采用非标记定量蛋白质组学方法,对3 种重楼属植物根部的蛋白质组进行分析,发现419 个差异表达蛋白质主要分布在代谢、细胞和单个有机体3 类过程,且不同重楼属植物皂苷含量高低与蛋白质的差异表达密切相关。上述研究为重楼生长培育的人工干预、相关药效成分的提高,以及物种的基因改良奠定了重要的理论基础。
4.3 基因克隆与功能研究
4.3.1 次生代谢产物相关基因 重楼属植物的主要有效成分为甾体皂苷,其通过甲羟戊酸途径合成,该途径涉及3-羟基-3-甲基戊二酰单酰辅酶A(3-hydroxy-3-methyl-glutaryl-CoA,HMG-CoA)、甲羟戊酸(mevalonic acid,MVA)和异戊烯基焦磷酸(isopentenyl pyrophosphate,IPP)等,相关基因见表2。研究发现HMGR-CoA 还原酶、鲨烯环氧酶(squalene epoxidase,SE)、鲨烯合酶(squalene synthase,SQS)、法尼基焦磷酸合酶(farnesylpyrophosphate synthase,FPS)是甲羟戊酸途径的主要关键酶。徐永艳等[56]对重楼属植物不同组织的HMGR基因的表达差异进行研究,结果表明其在根中的表达量最高,与刘军等[57]的结果一致。为进一步研究重楼皂苷合成机制,多位学者以滇重楼为材料,分别克隆获得SE、FPS 和SQS 基因,还发现重组SQS 蛋白可在大肠杆菌中表达[58-61]。这些研究为探讨滇重楼皂苷的合成生物学研究提供了科学依,对用基因工程手段来调节重楼属植物皂苷代谢途径中关键酶的表达,具有重要的意义及应用价据值。此外,众多学者以滇重楼、多叶重楼为材料,成功分离克隆获得了多个调控重楼皂苷生物合成的基因,包括甲羟戊酸焦磷酸脱羧酶(mevalonate pyrophosphate decarboxylase,MPD)、环阿屯醇合酶(cycloartenol synthase,CAS)、固醇-C24-甲基转移酶(sterol-C24-methyl transferase,SMT)等[58-64],为研究重楼属植物皂苷合成的作用机制和表达调控奠定基础。

表2 重楼属植物功能基因克隆情况Table 2 Cloning of functional genes in genus Paris
除上述相关功能基因研究外,在重楼属有效成分甾体皂苷生物合成途径中,糖基转移酶及细胞色素P450 酶、异戊烯焦磷酸异构酶和鲨烯合成酶基因也被发掘克隆。董栩成功克隆出滇重楼植物的糖基转移酶基因及细胞色素P450 基因的全长序列,发现在不同组织中基因表达量有差异。江雪梅等[65-66]采用RT-PCR 方法扩增了功能基因IPPI、SS 的部分序列,并分析它们在根、茎、叶3 个组织中的表达差异,也发现在不同组织部位中基因的表达模式不同,这些研究为探讨今后甾体皂苷合成的机制提供科学参考。
4.3.2 参与休眠解除和菌根互作相关的基因 赤霉素(gibberellin,GA)是一种促进生长的植物激素,可以调节种子发育,促进种子萌发,与植物种子休眠关系密切。滇重楼中赤霉素合成代谢途径的关键酶GA 氧化酶基因(GAox)已被成功克隆出来,GAox基因在滇重楼的不同器官中的表达,以及其种子的不同萌发阶段具有很明显的差异[67-69],且NCED、CYP707A、ABI2、GA20ox2、GA20ox3基因参与滇重楼种子休眠解除过程,是滇重楼种子休眠过程的重要基因。此外,陈瑶等[70]应用高通量转录组测序技术,分析了华重楼种子萌发前后激素信号转导途径和合成通路中的差异表达基因,发现GA 和IAA 组合能促进种子萌发,其调控机制可能与GA、IAA 合成代谢和信号转导相关。为探究重楼属植物的种子共生萌发分子机制,张华等[71]研究了接种不同外源性从枝菌根(arbuscular mycorrhizae,AM)真菌对滇重楼幼苗基因表达的影响,结合AM 真菌对种子萌发及幼苗化学成分的相关研究,推测薄壁两性囊霉和崔氏原囊霉菌株,可望作为培育滇重楼菌根化苗的理想菌株。上述基因的相关研究为滇重楼可持续利用提供了重要参考,然而,这些基因的功能远未阐明,有待进一步深入。
4.3.3 其他基因 在滇重楼的生长发育中,溶性无机焦磷酸酶(soluble inorganic pyrophosphatase,sPPase)基因可调控其休眠和萌发、光能利用等途径。赵爽等[72]从滇重楼中克隆到PpsPPase基因,初步实现了PpsPPase在原核细胞中的表达,可为进一步研究PpsPPase在滇重楼生长发育过程中的功能提供信息和依据。
5 结语与展望
随着分子生物学技术的不断发展,新一代高通量测序技术的开发,有力地推动了重楼属植物的分子生物学研究。然而,由于重楼属树种自身特性和地理分布,加上其分子生物学研究团队力量薄弱,资金投入不足等原因,分子生物技术在重楼属植物中的应用仍存在诸多难点。目前在重楼属植物中,已开发多种分子标记,用于其遗传多样性及遗传指纹图谱研究;同时基于DNA 分子片段的物种鉴别、分类及系统进化研究也取得了一些进展,但该属植物的分类学和系统学依然存在争议。此外,利用转录组学、蛋白组学和基因克隆技术对重楼属植物的功能基因研究也取得了一定进展,目前虽已克隆获得多个相关功能基因,但主要集中在滇重楼的次生代谢产物重楼皂苷合成、菌根互作和休眠解除相关基因方面,对重楼属其他物种的相关功能基因研究还知之甚少,且对环境胁迫的品质形成机制等仍存在一定局限性。当前众多学者虽然克隆获得了多个重楼属植物生物合成相关基因,但是,围绕这些基因开展的基因调控工作却很少,其生物合成途径中还有很多未解之谜,有待进一步深入发掘,为相关优良种质培育和资源的合理利用奠定基础;同时,由于重楼属植物拥有目前科学界已知的最大基因组,是研究基因组大小变化的理想模型,破译重楼属植物基因组对研究基因组大小进化以及重楼皂苷生物合成通路具有重要意义。今后对重楼属植物的研究中,应充分整合基因组学、转录组学、蛋白组学、代谢组学等多组学的技术手段,加快对重楼属植物功能基因的探索,为重楼属植物的进一步开发和利用提供理论依据。
利益冲突所有作者均声明不存在利益冲突
猜你喜欢
今日农业(2021年11期)2021-08-13
红楼梦学刊(2020年2期)2020-02-06
当代陕西(2019年6期)2019-04-17
中成药(2018年9期)2018-10-09
中成药(2017年9期)2017-12-19
中成药(2017年7期)2017-11-22
中成药(2017年8期)2017-11-22
中成药(2017年6期)2017-06-13
中国药业(2014年16期)2014-05-14
遗传(2014年3期)2014-02-28
