
梵净山国家级自然保护区叶蝉群落结构特征
2021-08-13 06:05陈晓晓高娅蓉熊康宁宋月华
环境昆虫学报 2021年4期
陈晓晓,高娅蓉,熊康宁,宋月华
(贵州师范大学喀斯特研究院/国家喀斯特石漠化防治工程技术研究中心,贵阳 550000)
叶蝉隶属于半翅目Hemiptera头喙亚目Auchenorrhyncha叶蝉科Cicadellidae,主要发生在森林和草地上,个体小,种类众多,以植物为食(Morris, 1971;郭慧芳,2011)。叶蝉是一类危害作物、草地、林木等植物的昆虫,常刺吸植物汁液,导致叶片出现淡白色斑点直至枯死,并且许多种类还传播植物病毒,给农林业带来严重的损失(廉振民和潘兴丽,2008;Roddeeetal., 2018),但其作为生态系统的重要组成部分,对物质循环、能量流动及环境变化监测等方面具有特殊的生态功能。相关研究表明,在不同区域环境、海拔和生境下昆虫群落组成和结构均有较大的差异,该差异反映了垂直带的特征和规律、植被演替变化以及环境干扰程度等,是重要的生态学评价指标之一(廉振民等,1997;曹凤麟等,2016;马玲等,2016)。
梵净山国家级自然保护区地处云贵高原向湘西丘陵的过渡地带,是多种植物区系地理成分汇集地,保护区内动植物种类多,生物多样性丰富,随着气候、地貌、海拔等自然环境的变化,呈现特有的分布和演化规律。叶蝉科昆虫是该区域生态系统的重要组成部分,对保护区内叶蝉物种进行调查研究,对不同垂直海拔林带和不同生境条件下的叶蝉类昆虫进行生物多样性统计,并且分析其分布特征,旨在探明该区域叶蝉群落结构特征及其与环境变化的关系,为保护区内生态环境质量监测提供参考依据,为叶蝉类昆虫在生物学、生态学及综合治理等研究上提供理论参考。
1 材料与方法
1.1 保护区自然概况
梵净山国家级自然保护区位于贵州省铜仁市江口县、松桃县和印江县三县的交界地带,地处108°45′50″-108°48′30″E、27°49′50″-28°1′30″N,地势显著隆起,是武陵山脉的最高峰,由于受到长期侵蚀,形成变质岩山脉地貌;属于中亚热带季风山地湿润气候,水热资源丰富,气候呈现垂直变化,并由下至上划分为中亚热带、北亚热带、南温带和中温带4个气候带(牛克锋,2014)。区内生境类型复杂,植被类型多样,植被呈现垂直带谱,主要有阔叶林、针叶林、竹林、灌丛和沼泽5个植被类型组,动植物种类丰富,是重要的生物资源基因库(黄威廉,2001)。
1.2 采样方法与标本处理
对2018年夏季(5-8月)梵净山保护区内的叶蝉科昆虫进行扫网采集,每月采集不同生境和林带上的叶蝉科昆虫,每个地方扫网50次,将采集后的标本(不包括若虫)保存在无水乙醇中,记录采集信息(时间、地点、高程、植被类型等),并依据叶蝉发生条件将其生存生境划分为以下5类:田地(耕种庄稼的土地)、草坡(多年生矮小草本植株密植的草地或坡地)、森林(具有高密度树木的群落)、林缘(在森林和草地或灌木群落之间的交错群落)和灌丛(矮小而丛生木本和草本植物的群落)(赵振强等,2015);依据海拔高度以及植被类型,研究区划分为4个垂直海拔林带:常绿阔叶林带(Ⅰ,<1 300 m)、常绿落叶阔叶混交林带(Ⅱ,1 300~1 900 m)、亚高山针阔混交林(Ⅲ,1 900~2 350 m)、亚高山灌丛草甸带(Ⅳ,2 350~2 570 m)(黄威廉,2001)。
标本鉴定主要依据《贵州大学馆藏昆虫(头喙类)模式标本图志》《中国斑叶蝉和塔叶蝉(半翅目:叶蝉科:小叶蝉亚科)》《中国竹子叶蝉》和《中国广头叶蝉(半翅目:叶蝉科)》等叶蝉分类著作及相关分类文献完成(李子忠等,1996,2006,2011,2014,2017;陈祥盛等,2012;宋月华等,2014;戴仁怀等,2018)。
1.3 数据分析与处理
1.3.1群落优势度分析
利用Excel 2010软件建立梵净山自然保护区叶蝉科昆虫分布信息数据库,以不同生境和垂直自然林带为单位,比较分析叶蝉群落组成与分布的差异,并分析类群的优势度。
1.3.2群落相似性分析
根据叶蝉各属在5种生境、4种垂直林带的分布数据,剔除仅在1个区分布的属,建立PAE矩阵,导入SPSS 22.0软件中,用最远相邻法,Jaccard系数为度量标准,进行层次聚类分析以及相似性分析。
1.3.3群落多样性分析
运用Shannon-Wiener多样性指数、Pieluo均匀性指数、Simpson优势度指数相结合来表示叶蝉群落的多样性(Simpson, 1949;Pielou, 1966;Peet, 1974)。
(1)Shannon-Wiener多样性指数
(1)
式中:H′为多样性指数;Pi=ni/N,ni为第i属的个体数,N为总个体数。
(2)Pieluo均匀性指数
J=H′/InS
(2)
式中H′为Shannon-Wiener多样性指数,S为属数。
(3)Simpson优势度指数
(3)
式中C为优势度指数,ni为第i种的个体数,N为总个体数。
2 结果与分析
2.1 叶蝉群落组成与分布
2.1.1不同生境条件下叶蝉群落组成与优势度
在保护区内5种不同生境类型中共捕获叶蝉标本4 279头,隶属于15亚科127属,叶蝉标本个体数依次为田地(1 483头,34.7%)>灌丛(1 246头,29.1%)>林缘(824头,19.3%)>草坡(491头,11.5%)>森林(235头,5.5%)。类群不同阶元数量顺序依次为灌丛(14亚科,85属)>林缘(13亚科,61属)>田地(9亚科,49属)>森林(12亚科,44属)>草坡(7亚科,33属)。统计分析结果表明(表1):在以不同生境条件为研究基础上,整个研究区内山小叶蝉属Salka和草叶蝉属Sorhoanus2属为优势类群,占总个体数的26.4%;柔突叶蝉属Abrus和干大叶蝉属Processina等27属为常见类群,占总个体数的51.6%;圆痕叶蝉属Japanagallia和广头叶蝉属Macropsis等98属为稀有类群,占总个体数的22.0%。在田地生境类型中,草叶蝉属Sorhoanus、山小叶蝉属Salka和尼氏叶蝉属Thapaia3属为优势类群,占该生境个体数的67.6%;二叉叶蝉属Macrosteles和吉岭叶蝉属Jilinga等12属为常见类群,占个体数的23.8%;条大叶蝉属Atkinsoniella和斑大叶蝉属Anatkina等34属为稀有类群,占个体数的8.5%。在草坡生境类型中,阿里山叶蝉属Alishania和干大叶蝉属Processina2属为优势类群,占该生境个体数的33.2%;广头叶蝉属Macropsis和横脊叶蝉属Evacanthus等12属为常见类群,占个体数的59.5%;条大叶蝉属Atkinsoniella和斑大叶蝉属Anatkina等19属为稀有类群,占个体数的7.3%。在森林生境类型中,条大叶蝉属Atkinsoniella为优势类群,占该生境个体数的13.2%;愈叶蝉属Maiestas和吉岭叶蝉属Jilinga等26属为常见类群,占个体数的77.4%;杨小叶蝉属Alnetoidia和泰小叶蝉属Thailocyba等17属为稀有类群,占个体数的9.4%。在林缘生境类型中,山小叶蝉属Salka和米小叶蝉属Mitjaevia2属为优势类群,占该生境个体数的30.5%;干大叶蝉属Processina和条大叶蝉属Atkinsoniella等17属为常见类群,占个体数的52.5%;网脉叶蝉属Dryodurgades和多脉叶蝉属Multinervis等42属为稀有类群,占个体数的17.0%。在灌丛生境类型中,叶蝉分布无优势类群;杨小叶蝉属Alnetoidia和优小叶蝉属Qadria等34属为常见类群,占该生境个体数的82.8%;凸唇叶蝉属Erragonalia和凹大叶蝉属Bothrogonia等51属为稀有类群,占个体数的17.2%。

表1 梵净山国家级自然保护区不同生境类型叶蝉的群落组成
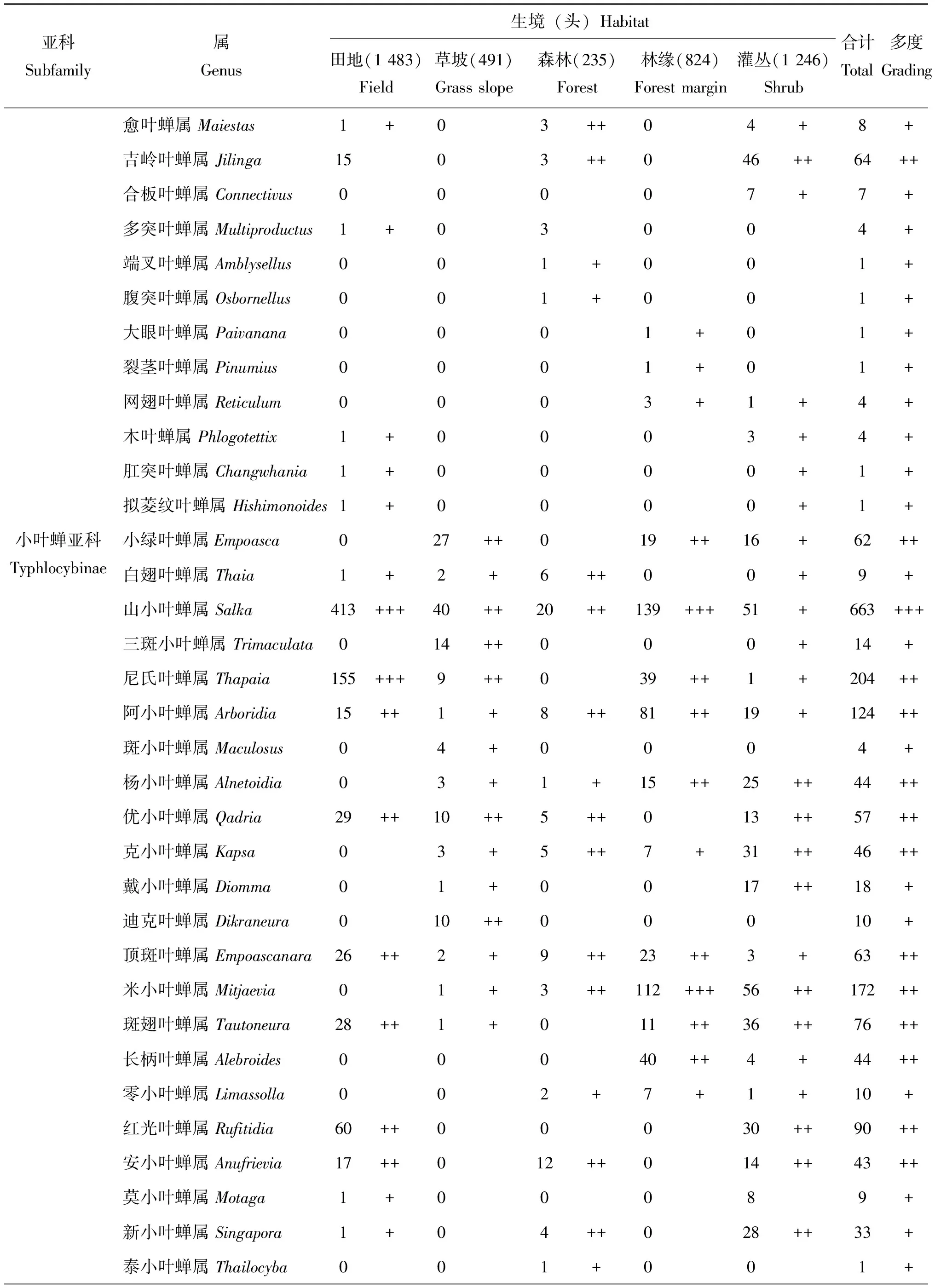
续表1 Continued table 1
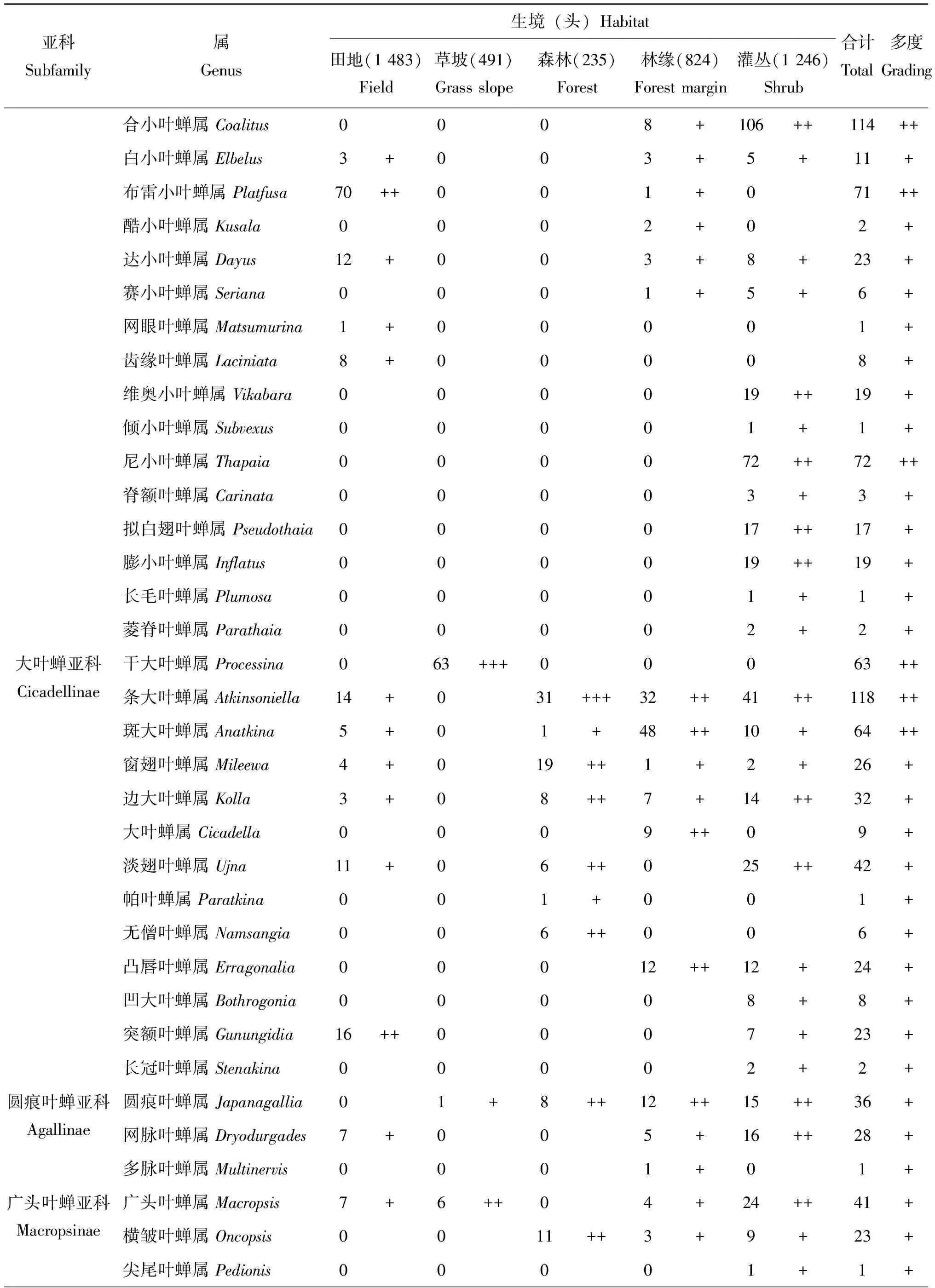
续表1 Continued table 1
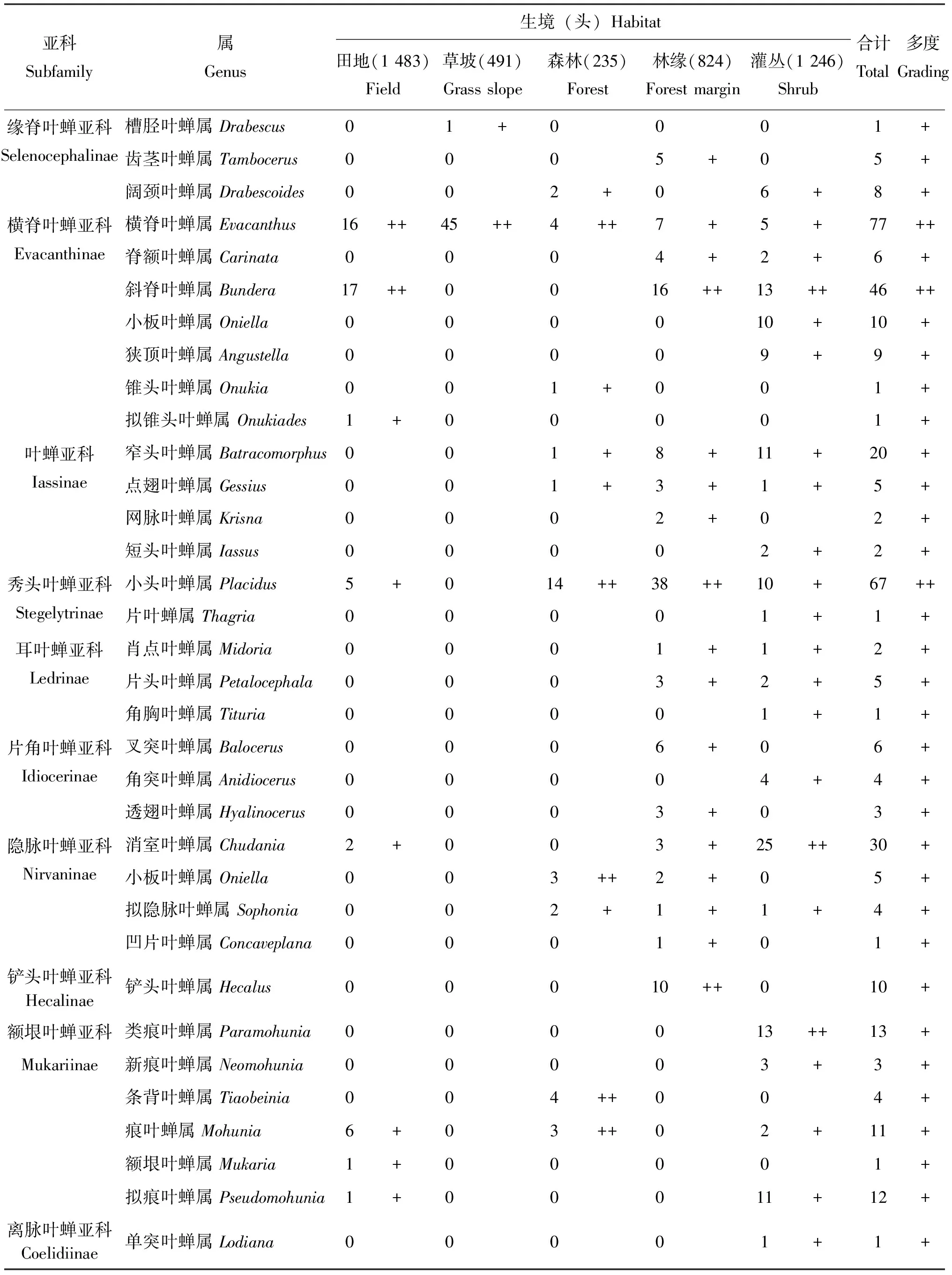
续表1 Continued table 1
2.1.2不同海拔垂直林带下叶蝉群落组成与优势度
在研究区内5种不同海拔垂直林带,共捕获叶蝉标本4 278头,隶属于15亚科127属,在不同海拔垂直林带中,叶蝉标本个体数依次为常绿阔叶林带(1 968头,46.0%)>亚高山灌丛草甸带(1 253头,29.3%)>亚高山针阔混交林带(838头,19.6%)>常绿落叶阔叶混交林带(219头,5.1%)。类群不同阶元数量顺序依次为亚高山灌丛草甸带(14亚科,88属)>常绿阔叶林带(10亚科,68属)>亚高山针阔混交林带(14亚科,66属)>常绿落叶阔叶混交林带(11亚科,39属)。统计分析结果表明(表2):在以不同海拔垂直林带环境为研究的基础上,整个研究区内山小叶蝉属Salka和草叶蝉属Sorhoanus2属为优势类群,占总个体数的26.1%;干大叶蝉属Processina和斜脊叶蝉属Bundera等27属为常见类群,占总个体数的51.7%;消室叶蝉属Chudania和小板叶蝉属Oniella等98属为稀有类群,占总个体数的22.2%。在常绿阔叶林带中,草叶蝉属Sorhoanus和山小叶蝉属Salka2属为优势类群,占该林带个体数的44.6%;顶斑叶蝉属Empoascanara和红光叶蝉属Rufitidia等15属为常见类群,占个体数的42.0%;窗翅叶蝉属Mileewa和边大叶蝉属Kolla等51属为稀有类群,占个体数的13.4%。在常绿落叶阔叶混交林带中,条大叶蝉属Atkinsoniella为优势类群,占该林带个体数的14.2%;愈叶蝉属Maiestas和吉岭叶蝉属Jilinga等23属为常见类群,占个体数的76.7%;窄头叶蝉属Batracomorphus和点翅叶蝉属Gessius等15属为稀有类群,占个体数的9.1%。在亚高山针阔混交林带中,山小叶蝉属Salka和米小叶蝉属Mitjaevia2属为优势类群,占该林带个体数的30.0%;缘毛叶蝉属Phlogothamnus和多室叶蝉属Subhimalus等17属为常见类群,占个体数的51.7%;肖点叶蝉属Midoria和片头叶蝉属Petalocephala等47属为稀有类群,占个体数的18.3%。在亚高山灌丛草甸带中,叶蝉分布无优势类群;圆痕叶蝉属Japanagallia和网脉叶蝉属Dryodurgades等34属为常见类群,占该林带个体数的82.4%;横脊叶蝉属Evacanthus和脊额叶蝉属Carinata等54属为稀有类群,占个体数的17.6%。
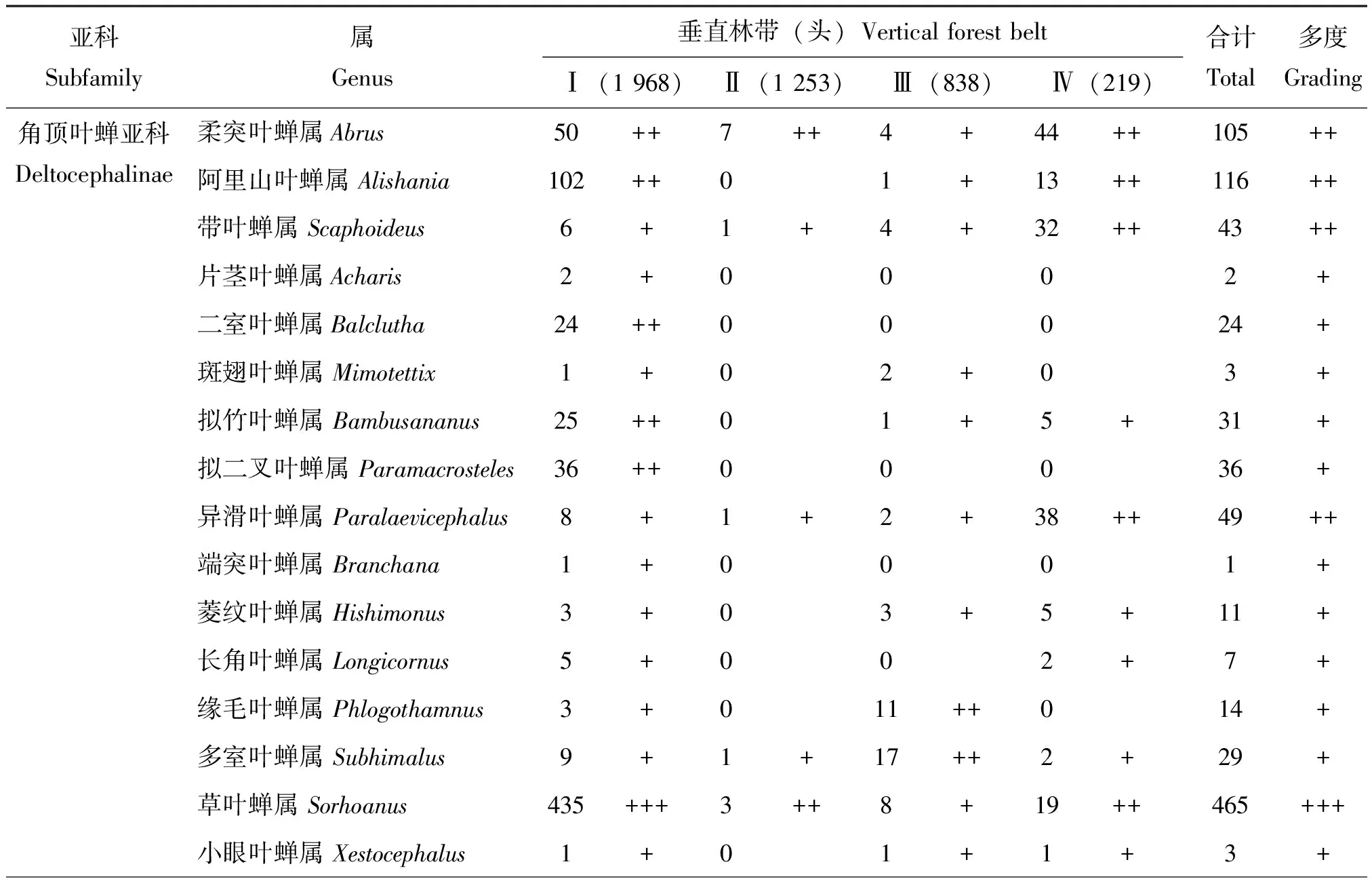
表2 梵净山国家级自然保护区不同海拔垂直林带叶蝉的群落组成
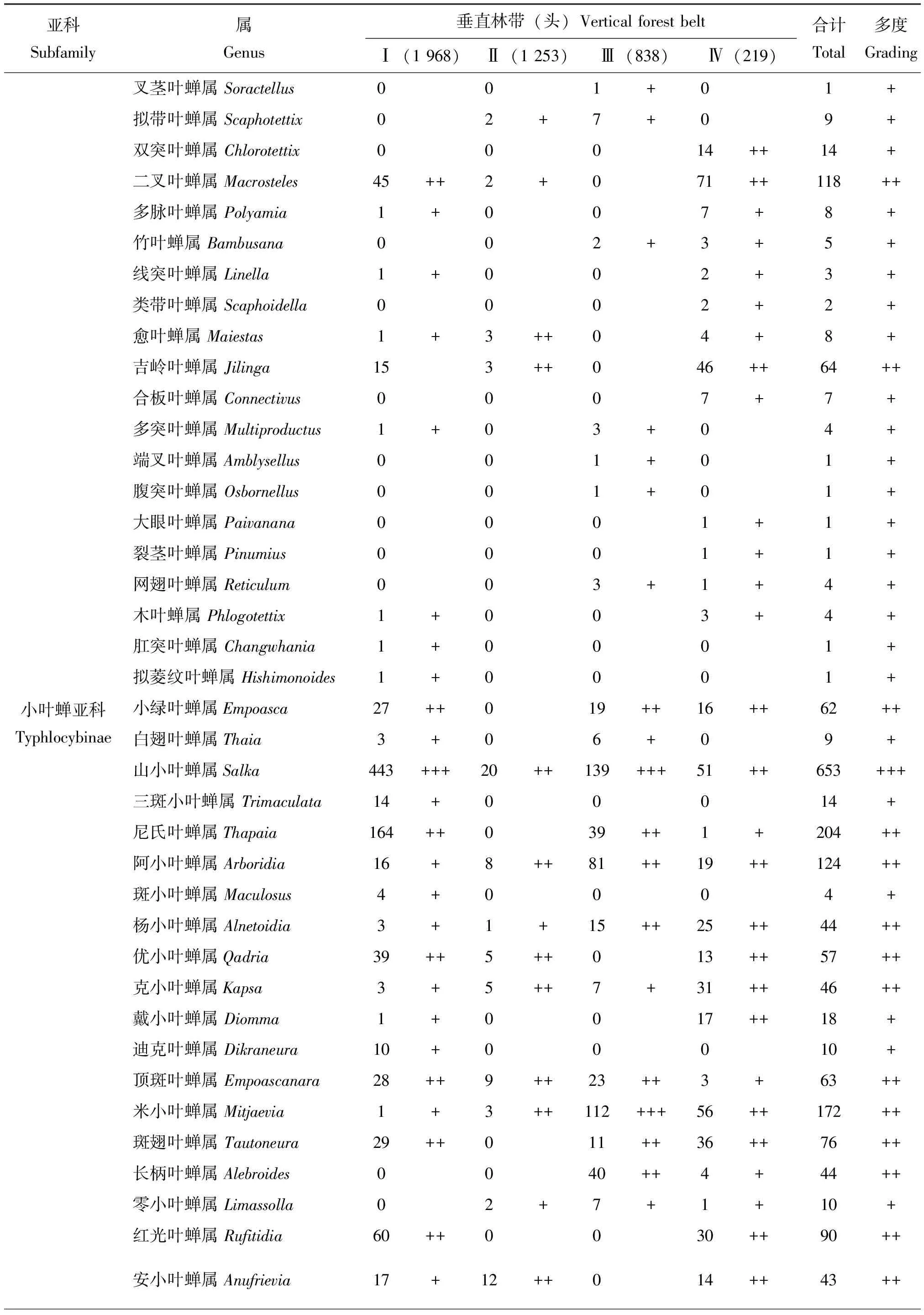
续表2 Continued table 2
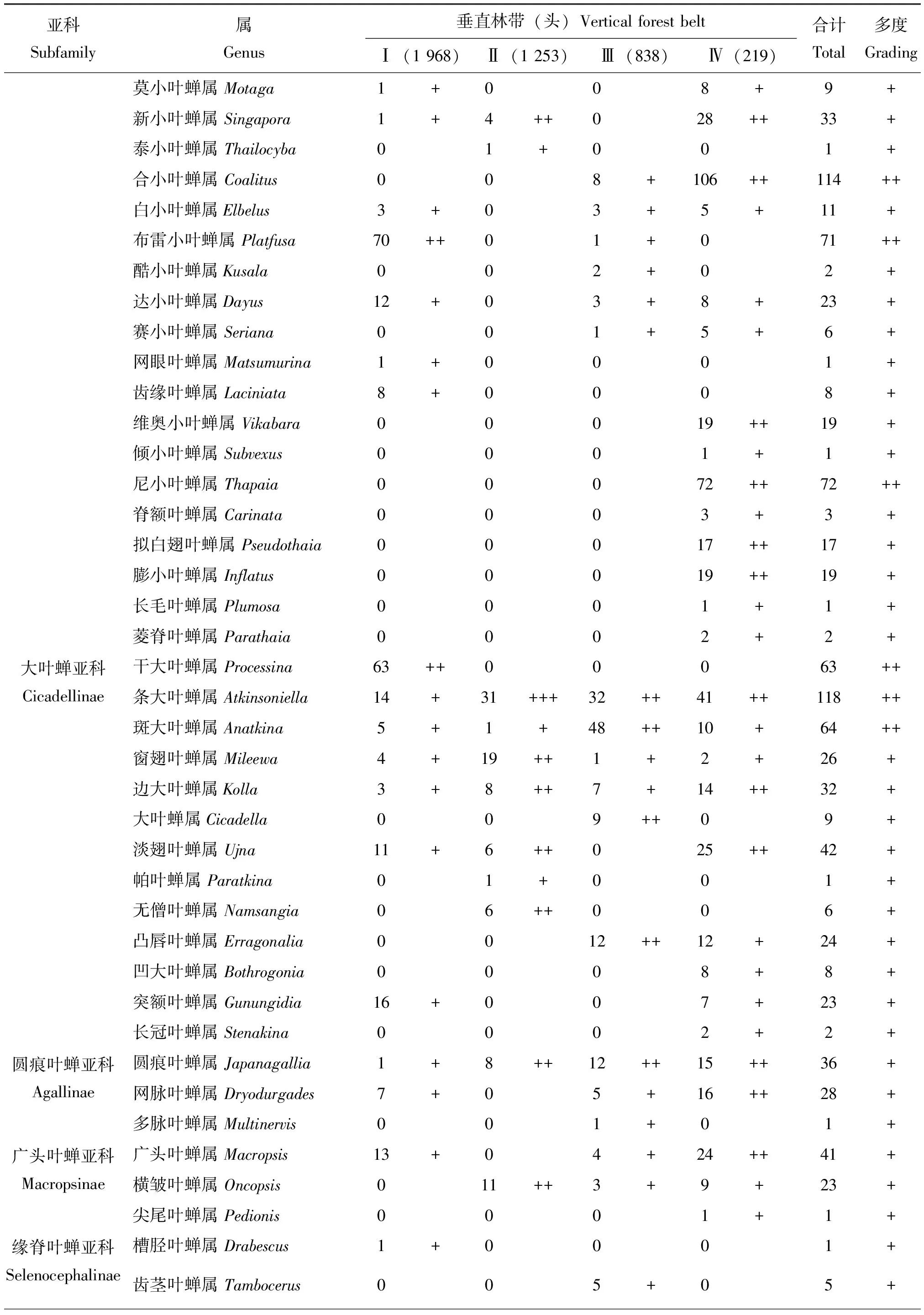
续表2 Continued table 2

续表2 Continued table 2
2.2 叶蝉类群的相似性
2.2.1不同生境叶蝉类群聚集性和相似性
为直观表明不同生境间叶蝉组成的相似性,使用层次聚类分析法对叶蝉类群组成的相似性关系进行分析(图1和表3)。结果表明,林缘和灌丛最为接近,首先聚为一团A,A和森林相聚后,组成聚类团B,B再与田地形成聚类团C,草坡与前面形成的大类群C最终相聚,组成整个聚类团D。其中,聚类团A内距离系数最小,灌丛和林缘之间约为1.5,相似性指数为1;森林和聚类团A的距离系数次之,约为13.5,相似性指数为0.596;田地和聚类团B的距离系数约为18.5,相似性指数为0.542,草坡与聚类团C的距离系数最大,为25.0,相似性指数为0.063。这表明生境的相似是叶蝉群落相似的基础,在自然条件下,灌丛和林缘最为相似,草坡与其他生境相比,其内部环境和结构更趋于简单化,生态系统脆弱,田地虽与其它生境截然不同,但其间多为退耕地,混有杂草、灌丛等植被。灌丛和林缘内叶蝉类群的相似性最高,物种交流最为频繁,生境之间关联性最强,而草坡与其他生境类型的物种交流程度最低。

图1 梵净山国家级自然保护区不同生境类型叶蝉类群聚类分析Fig.1 Cluster analysis of leafhopper groups at different habitats in Fanjingshan National Nature Reserve

表3 梵净山国家级自然保护区不同生境类型叶蝉类群相似性矩阵
2.2.2不同海拔垂直林带叶蝉类群聚集性和相似性
对不同海拔垂直林带间叶蝉类群组成的相似性关系进行分析(图2和表4)。结果表明,常绿阔叶林带和亚高山灌丛草甸带最为接近,首先聚为一团a,a和亚高山针阔混交林带相聚后,组成聚类团b,b再与常绿落叶阔叶混交林带组成整个聚类团c。其中,聚类团a内距离系数最小,常绿阔叶林带和亚高山灌丛草甸带之间约为1.5,相似性指数为1;亚高山针阔混交林带和聚类团a的距离系数次之,约为16.5,相似性指数为0.166;常绿落叶阔叶混交林与聚类团b的距离系数最大,为25.0,相似性指数为0.12。这表明不同林带环境的相似与否是叶蝉群落相似性大小的基础,在自然条件下,叶蝉具有趋光性,常绿阔叶林带和亚高山灌丛草甸带内食物条件和透光条件最为相似,常绿落叶阔叶混交林与其他林带相比,生态系统脆弱,易受人为干扰。常绿阔叶林带和亚高山灌丛草甸带的叶蝉类群相似性最高,物种交流最为频繁,林带之间关联性最强,而常绿落叶阔叶混交林带与其林带的物种交流程度最低。

图2 梵净山国家级自然保护区不同垂直林带叶蝉类群聚类分析Fig.2 Cluster analysis of leafhopper groups at different forest belts in Fanjingshan National Nature Reserve

表4 梵净山国家级自然保护区不同海拔垂直林带叶蝉类群相似性矩阵
2.3 叶蝉群落多样性
2.3.1不同生境叶蝉群落多样性
多样性计算结果显示(表5):各生境H′指数空间分布为:灌丛>森林>林缘>草坡>田地;J指数空间分布为:森林>灌丛>草坡>林缘>田地;C指数空间分布为:田地>草坡>林缘>森林>灌丛。结果表明,叶蝉群落组成在灌丛最为丰富,且各类群的数量也相对均匀,因此其多样性指数最高。森林和林缘的多样性指数相近,但均匀性前者较高,与后者差异显著。田地的优势度指数最高,说明其群落内物种分布较不均匀,自然环境与其他生境存在差异,个体数虽最大,但优势类群突出,多样性指数相对较低。研究结果表明,生境条件不同,叶蝉群落在不同生境间的分布有较大差异,从田地到灌丛,多样性指数和均匀度指数逐渐变大,而优势度指数逐渐减小。

表5 梵净山国家级自然保护区不同生境类型叶蝉群落多样性
2.3.2不同垂直海拔林带叶蝉群落多样性
多样性计算结果显示(表6):各林带H′指数空间分布为:亚高山灌丛草甸带>亚高山针阔混交林带>常绿落叶阔叶混交林带>常绿阔叶林带;J指数空间分布为:常绿落叶阔叶混交林带>亚高山灌丛草甸带>亚高山针阔混交林>常绿阔叶林带;C指数空间分布为:常绿阔叶林带>亚高山针阔混交林带>绿落叶阔叶混交林带>亚高山灌丛草甸带。叶蝉类群组成在亚高山灌丛草甸带最为丰富,且各类群的数量也相对均匀,因此其多样性指数最高。常绿落叶阔叶混交林带和亚高山针阔混交林带的多样性指数相近,但均匀性前者较高,与后者有一定差异。常绿阔叶林带的优势度指数最高,优势类群突出,说明其群落内物种分布较不均匀,个体数虽然最大,但多样性指数相对较低。研究结果表明,随着海拔的升高及植被带的变化,叶蝉类群分布在不同林带间有较大差异,而且优势度指数逐渐减小。

表6 梵净山国家级自然保护区不同海拔垂直林带叶蝉群落多样性
3 结论与讨论
昆虫群落的组成与结构是研究昆虫群落特征变化的重要指标之一。许多研究表明,昆虫群落特征的变化,是昆虫与植物长期作用的结果,植被组成越丰富,昆虫群落越多样(蒋杰贤等,2011;李丽丽等,2011)。本研究中,梵净山国家级自然保护区叶蝉群落组成丰富,仅本次初步调查就有15亚科,127属,并且随着植被组成变化和海拔升高,叶蝉类群在5种生境和4种林带间的分布存在较大差异,在植被组成丰富、光照条件充足的环境中,叶蝉类群多样性增加;在植被类型单一且密度大的环境下,叶蝉类群数量趋于减少。在不同生境下,灌丛内植被组成丰富,多为矮小而丛生的木本和草本植物,因此分布的叶蝉类群数量最多,物种的丰富度最大;而在草坡内,由于内部环境单一,坡度较大,多为矮小草本植株密植的草地或坡地,所以分布的叶蝉类群数量最少;在不同垂直林带中,亚高山灌丛草甸带内多为低矮植物,以杜鹃—大箭竹灌丛为主,区内光照条件较好,叶蝉分布数量最多,多样性较大;而在常绿落叶阔叶混交林带内,常绿植物较高,随着海拔增高,落叶植物增加(沈定荣等,1985),林间密度大,分布的叶蝉类群最少。除了自然因素导致的群落结构变化,人为因素也产生一定程度的影响。田地中,叶蝉个体数虽然最多,但易受人类活动干扰,其间多为退耕地,混有杂草、灌丛等植被,标本采集更为便利,而植被组成较为单一,境内生态系统不稳定,所以分布类群种类较少。常绿落叶阔叶混交林带处于梵净山旅游景点区域,该海拔梯度的人为干扰尤为明显(夏常英等,2020),旅游设施和旅游过程带来的植被破坏、林地减少是不可避免的,因此该林地个体数和类群数均最少。虽然自然环境差异导致群落多样性呈现不同程度的变化,但是群落结构中优势类群、常见类群、稀有类群在整体上差异甚微,无论是以生境还是林带为研究基础,整个研究区内山小叶蝉属Salka和草叶蝉属Sorhoanus均为梵净山叶蝉分布的优势类群,横脊叶蝉属Evacanthus、顶斑叶蝉属Empoascanara、阿小叶蝉属Arboridia、山小叶蝉属Salka、异滑叶蝉属Paralaevicephalu、带叶蝉属Scaphoideus均有分布。
自然环境的相似是叶蝉群落相似的基础,区间自然环境相似性越高,叶蝉群落越相似,不同生境和不同林带中存在的差异,导致了叶蝉群落相似性的不同(刘高峰和杨茂发,2003)。在不同生境条件下,灌丛和林缘在植被组成和植被密度、小气候、光照等方面最为相似,其叶蝉类群组成相似性最高,生境之间关联性最强;草坡与森林内部自然环境差距明显,其中草坡内部自然环境简单,多为矮小草本植株,光照充足,容易受人类活动的影响,而森林内多为高密度树木,光照较低,生境内环境相对稳定,因此,以上二生境叶蝉群落组成相似性最低,区间关联性弱。在不同林带下,常绿阔叶林带和亚高山灌丛草甸带相似性最高,群落差异不大,两者在植被类型(草被发达)、光照条件等方面具有一定的相似性,但在植被群落类型多样化、温度、湿度等其它自然环境上差异显著。常绿阔叶林带中树种组成复杂,常绿植物多,温度较高,而亚高山灌丛草甸带植被组成较简单,以灌丛为主,随着海拔上升,温度降低。形成这种现象的原因可能是由于常绿落叶阔叶混交林带和亚高山针阔混交林带在一定程度上受人为活动的影响,其内部自然生态环境遭到破坏,原生植被逐渐减少,导致叶蝉类群多样性减少,而常绿阔叶林带和亚高山灌丛草甸带原生植被覆盖较高,所以叶蝉类群相对丰富,两者相似性高。
昆虫群落分布受植被种类和密度的直接影响。叶蝉以刺吸植物汁液为食,其发生条件与植物密切相关,植食性昆虫的多样性和植物多样性呈正相关,而光照、小气候、养分都会受到生境和林带结构层次变化的影响,进而影响植被组成的多样性(Barbleretal., 2008;Takietal., 2010)。一般情况下,昆虫群落的多样性对昆虫群落的系统稳定性有明显影响,群落多样性越高,内部关系越复杂,群落环境越稳定,抗干扰的能力越强,群落的均匀度指数就高(刘德广等,2001;张淑莲等,2005)。该研究表明,随着生境条件和林带环境的不同,海拔的升高和植被带的变化,叶蝉类群的分布存在较大差异,多样性指数随着均匀度指数的增大而增大,而优势度指数则相反。从田地到灌丛以及从常绿阔叶林带到亚高山灌丛草甸带,多样性指数逐渐增大,叶蝉类群组成在灌丛和亚高山灌丛草甸带最为丰富,且各类群的数量也相对均匀,而田地和常绿阔叶林带的优势度指数最高,说明其群落内物种分布较不均匀,个体数虽最大,但优势类群突出,多样性指数相对较低。
研究以梵净山国家级自然保护区内叶蝉科昆虫为研究对象,首次调查了保护区内叶蝉群落的组成和分布,阐述了不同生境和不同垂直林带中叶蝉群落结构的相似性和多样性,叶蝉群落结构的差异反映了人为活动对物种多样性的影响,为梵净山生态环境保护提供了一定参考依据,也为梵净山其它昆虫类群的多样性研究奠定了基础,丰富了梵净山昆虫物种多样性的数据,同时该研究对理解我国区域性昆虫物种多样性的特征、动态变化和形成机制具有一定的理论意义。
致谢:感谢袁周伟、谭超、苑晓伟在野外标本采集及分类过程中给予的帮助。
猜你喜欢
水土保持学报(2022年5期)2022-10-10
西北农林科技大学学报(自然科学版)(2022年3期)2022-04-01
农业科技与信息(2020年14期)2020-12-18
农业工程学报(2020年11期)2020-07-22
安徽农学通报(2020年6期)2020-04-17
绿色科技(2019年14期)2019-11-19
江苏农业科学(2019年5期)2019-09-02
绿色科技(2019年13期)2019-08-31
陕西农业科学(2019年6期)2019-07-19
浙江林业(2017年8期)2017-11-13
