
CRISPR基因编辑技术及其在眼科疾病中的应用进展
2021-08-23 09:32马丹宝钟玮晔综述于晨审校
眼科学报 2021年7期
马丹宝,钟玮晔 综述 于晨 审校
(1.深圳湾实验室肿瘤研究所,广东 深圳 518132;2.斯特拉斯堡大学生命科学学院,法国 斯特拉斯堡)
基因编辑是通过导入、修改、沉默特定基因,以研究基因功能,构建疾病模型,以及治疗特定疾病的一种新兴技术[1]。CRISPR(clustered regularly interspaced short palindromic repeats)技术的发明,大大提高了基因编辑的灵活性,并加快了基因编辑技术从实验室走向临床治疗的步伐[2]。眼球独特的生理结构,使眼科疾病在基因编辑疗法的应用上具有先天的优势。眼球体积较小,需要的治疗载体较少。眼部区域具有免疫赦免,这既可避免外源性物质引起的炎症和免疫反应,也有利于基因编辑载体维持其活性。同时,慢病毒和AAV等载体介导的眼科疾病基因疗法,不会引起全身性反应[3]。另外,多种严重眼科疾病与基因突变密切相关。这些特点使眼科疾病的治疗对基因编辑疗法有着强烈的需求,为其在眼科疾病治疗中的率先应用创造了条件。本文将从CRISPR/Cas9(Cas9,CRISPR associated protein 9,曾使用Cas5、Csn1、Csx12等名称)基因编辑技术的发展、基因编辑疗法在眼科疾病治疗中应用的现状、以及CRISPR基因编辑技术的最新发展等几个方面,介绍CRISPR基因编辑技术及其在眼科疾病治疗中应用的前沿进展。基因疗法的载体及递送策略,相关综述文献已有阐述[4],在此将不再赘述。
1 第1代CRISPR-Cas9基因编辑技术
CRISPR系统是一种原核生物获得性免疫系统[5-6]。它由一系列CRISPR相关蛋白、可编程RNA分子和Cas9核酸内切酶所组成。当外源病原体侵入细菌后,该系统将外源DNA片段整合到细菌基因组中,并以该片段为模板转录加工出CRISPR RNA(crRNA)[7]。病原体再次感染细胞时,该crRNA介导Cas9蛋白靶向切割病原体DNA,从而达到消除病原体,保护细胞的作用。
2012年,Jennifer Doudna和Emmanuelle Charpenier将CRISPR/Cas9系统改造,实现了由引导RNA(guide RNA)介导的DNA靶向切割[2]。该系统由Cas9核酸内切酶和单链引导RNA(single-stranded guide RNA,sgRNA)所组成(图1)。SgRNA引导Cas9特异结合与sgRNA上一段RNA序列互补的DNA序列,随后在附近将DNA双链切割。被切割后的DNA形成双链断裂(double strand break,DSBs),DSBs会激活内源性DNA损伤修复机制[8]。例如非同源性末端接合(nonhomologous end-joining,NHEJ)在将DSBs重新连接的过程中,会产生插入或删除突变(indel),而indel突变往往造成编码位移(frameshift)以及基因敲除[9-10]。如用多个sgRNA介导多处DSBs的形成,则会促进DSBs之间大片段的染色质重排(genomic rearrangement),如染色体内易位(intrachromosomal translocation)和染色体内倒位(intrachromosomal inversions)[11-13]。这些机制均可实现高效的基因敲除。
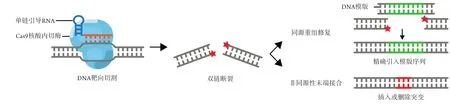
图1 第1代CRISPR/Cas9基因编辑技术Figure 1 The first generation of CRISPR/Cas9 gene editing technology
除NHEJ机制外,CRISPR/Cas9切割导致的DSBs还可诱导另一种DNA损伤修复机制——同源重组修复(homology-directed repair,HDR)[14]。在单链DNA分子或双链质粒DNA模板存在的情况下,HDR可精确引入模板序列,从而导致基因突变、基因修正、基因插入[15]。
2 CRISPR-Cas9基因编辑疗法治疗眼科疾病的最新进展
2.1 Leber先天性黑矇(已在患者中开展I期临床试验)
一部分10型Leber先天性黑矇(Leber congenital amaurosis 10,LCA10)是由CEP290基因的功能缺失性突变所导致[16],绝大多数患者出生后视力低下或完全失明。在高加索人种中,最常见的LCA10是由CEP290基因上的IVS26突变所导致[17-18]。该突变位于26号内含子,突变创造了一个新的剪接位点,导致mRNA被错误剪接及蛋白翻译的提前终止[19]。同时,由于CEP290基因较大(7.5 kb),受载体容量限制,传统依赖于AAV载体引入全长基因的方法无法对其实现修复。CRISPR基因编辑治疗旗舰公司Editas开发了一款使用CRISPR-Cas9的基因编辑产品EDIT-101(AGN-151587)[20],该产品在IVS26突变两侧切割DNA,造成IVS26突变的删除,从而诱导正常的mRNA剪接,并恢复CEP290功能。目前美国食药监局FDA已经批准该疗法的临床试验,2020年,一位先天失明患者成为该疗法的第1位临床试验患者,另外17位成人及儿童患者随后将接受EDIT-101单眼的视网膜下注射[21]。
2.2 2A型Usher 综合征(已完成动物上的临床前研究,即将开展I期临床试验)
2A型Usher综合征(Usher syndrome 2A),由USH2A基因上的突变所导致[22],其特征为听力损伤和由视网膜色素变性(retinitis pigmentosa,RP)引起的进行性视力丧失。USH2A基因编码一个位于感光细胞连接鞭毛(connecting cilium)处的跨膜基质蛋白,该蛋白被认为对于稳定感光细胞连接处外侧结构具有重要作用[23]。该基因13号外显子上c.2299delG突变导致编码位移及蛋白产物缺损,引起鞭毛缺陷。Editas公司的基因编辑产品EDIT-102利用Cas9在13号外显子两侧进行切割,引起13号外显子的敲除[24]。缺失13号外显子的USH2A基因可产生功能正常的蛋白。该策略已在小鼠中被证明有效,目前正在准备开展临床试验。
2.3 单纯疱疹病毒性角膜炎(已完成动物上的临床前研究,即将开展I期临床试验)
单纯疱疹病毒性角膜炎(herpes simplex keratitis,HSK)是单纯疱疹病毒(herpes simplex virus,HSV)引起的一种严重感染性角膜疾病[25]。其中,复发性HSV感染是感染性角膜失明的主要原因。复发性HSK由潜伏于三叉神经节处的HSV复发,并随后引起基质的炎症反应所导致[26]。CRISPR/Cas9系统可以特异性在病毒基因组导入突变,是一种新型的抗病毒策略。为寻找抗病毒的最佳靶点,Editas的研究人员[27]对HSV基因组进行了筛选,他们发现UL48和RL2是两个最为有效的靶点。对这两个靶点的突变高效抑制HSV的复制。动物实验[27]表明:靶向突变UL48和RL2可以减少91%的角膜损伤,以及泪膜中75%的病毒载量。这种基于CRISPR/Cas9的抗病毒策略有望成为对抗复发性HSV感染的新一代治疗方法。
2.4 视网膜色素变性(正在动物上进行临床前研究)
视网膜色素变性是一类以进行性感光细胞和色素上皮细胞功能障碍为特征的遗传性疾病,是世界范围内常见的致盲性眼病[28]。RP在人群中发病率约为1/3 000~1/7 000,RP可通过常染色体显形、隐形或X染色体关联方式遗传。RP的发病机制尚不完全明确,目前已有几十个基因的突变被认为与RP高度相关,目前的研究热点包括RHO、PRPF31、RP1、PRPH2、IMPDH1、NR2E3、SNRPN200、CRX等基因[29]。针对这些基因突变,科学家已经开始设计相关的基因编辑治疗策略。例如,RHO基因编码1个感光细胞蛋白Rhodopsin,在视觉信号转导中有重要作用[30]。RHO一个基因拷贝上的P23H突变,导致Rhodopsin蛋白的错误折叠和蛋白降解途径的过载,引起了野生型RHO拷贝蛋白产物的降解[31]。Giannelli等[32]的研究表明:在小鼠模型中,通过CRISPR/Cas9系统靶向P23H突变的RHO基因拷贝,可以特异性敲除该基因拷贝,减缓Rhodopsin的降解并促进视力的恢复。
3 新一代CRISPR基因编辑技
第1代CRISPR-Cas9基因编辑技术,如依赖于HDR或NHEJ的基因编辑,均以Cas9等CRISPR相关核酸酶对DNA的靶向切割,以及随后产生的DSBs为前提。虽然这一途径极大加速了基因编辑的过程,但随着DSBs的修复,NHEJ通路介导的indel突变难以避免[33-34]。这一根本性缺陷,有可能影响基因编辑疗法的安全性及治疗效果。另外,HDR介导的精确基因编辑,只发生在G2和S期[35-36],理论上只可应用于增殖活跃的细胞,很难被应用于眼科疾病中。为解决上述限制,科学界正积极开发新一代CRISPR基因编辑系统,包括单碱基编辑(base editors),反转率相关CRISPR编辑(prime editors)和转座子相关CRISPR编辑(transposon-associated CRISPR)等。这些新的系统,均有望被开发为新一代基因编辑疗法,应用于眼科疾病。
3.1 单碱基编辑
单碱基编辑是一种不经DSBs介导而引入点突变的技术,以保留Cas9蛋白单链切割能力的nCas9为基础,融合腺核苷去氨酶(Adenosine deaminase),介导靶向的去氨基反应[37-38]。目前已有多种单碱基编辑系统面世,包括CBEs(介导C到T编辑)[37-38]、ABEs(介导A到G编辑)[39]、GBEs(glycosylase base editors,介导C到G编辑)[40]等。单碱基编辑器(以CBEs为例)首先对靶向DNA单链进行切割,随后系统中的去氨酶融合到切割部位,催化去氨基反应。去氨基反应将C转变为U,导致U-G错配。U-G错配随后会被内源DNA修复机制修改为T-A,完成C-T编辑。其他单碱基编辑系统,根据融合酶的不同,作用机制有所不同。但总体来说,单碱基编辑既不依赖于细胞周期[41],也不引入DSBs,有望被广泛应用于眼科多种疾病的治疗。
3.2 反转录相关CRISPR编辑
与单碱基编辑相似,反转录相关CRISPR编辑系统仍然采用nCas9[42]。不同的是,反转录相关CRISPR编辑利用反转录酶结构域与nCas9融合。同时,反转录相关CRISPR编辑采用pegRNA(prime editing guide RNA)代替sgRNA,其上除了含有靶向DNA的引导RNA序列外,还含有反转录编辑的模板。基因编辑时,nCas9首先对靶向区域单链进行切割,随后切割开的3’端DNA与位于pegRNA上的互补序列结合,并在反转录酶结构域的作用下,以pegRNA为模板合成互补DNA序列。随后,这一新合成的DNA序列被细胞内源DNA修复机制整合入双链DNA中,完成基因编辑。相比单碱基编辑,反转录相关CRISPR编辑可以引入任意的DNA突变,具有更大的灵活性。但同时,这一系统有一定的概率引入indel突变副产物。尽管如此,反转录相关CRISPR编辑由于具有较自由的插入和删除特定基因片段的能力,仍被认为具有巨大的临床应用前景。
3.3 转座子相关CRISPR系统
除上述系统外,还有2个研究小组[43-44]独立开发了转座子相关的CRISPR系统。以Strecker等[43]开发的CAST系统为例,该系统由3个部分组成:CRISPR/Cas12k靶向系统,TnsB-TnsC-TniQ转座子系统,包含两侧转座子序列的DNA模板。该系统进行基因编辑时,Cas12k靶向特异DNA序列,TnsB和TnsC将模板序列切割加工,TniQ将加工后的模板插入靶细胞基因组DNA中,完成基因编辑。这类转座子相关的CRISPR基因编辑系统完全独立于靶细胞内源DNA损伤修复机制,理论上可大大提高基因编辑的效率、提高可插入片段的长度,适合大片段的基因插入和敲除等操作。但与此同时,转座子末端序列会整合入受体细胞基因组,这使得基因编辑不可避免地引入了额外的序列。同时该类系统目前还未应用于真核细胞。尽管如此,类似的工具被改造并适用于真核细胞后,也可成为眼科疾病基因编辑疗法的有力工具。
4 结语
基于CRISPR的基因编辑疗法,是一种从根本上改变疾病治疗方案的技术。目前该疗法仍存在递送效率低、存在脱靶效应等缺陷。但总体来看,通过CRISPR基因编辑实现眼科疾病的治疗,具有很大的潜力,为多种眼科疾病、特别是罕见病的治疗带来了曙光。以单碱基编辑为代表的新一代CRISPR基因编辑工具,未来有望被开发为安全性更高、治疗效果更好的眼科疾病基因疗法。例如一些由基因点突变引起的眼科疾病,如PRPH2基因上多种点突变引起的视网膜色素变性[45-46]、LCAT基因上IVS4点突变引起的鱼眼病(fish-eye disease)[47]等,可采用单碱基编辑器加以修复。随着Editas公司LCA10临床试验的展开,以及多种眼科疾病的治疗方案完成临床前实验,CRISPR基因编辑对眼科疾病治疗的研究也即将进入爆发阶段。在不久的将来,人类必然可以通过改变患者的基因组实现对多种眼科疾病的治疗。
猜你喜欢
中华实验眼科杂志(2022年3期)2022-11-14
中老年保健(2022年1期)2022-08-17
广西医科大学学报(2022年6期)2022-07-13
保健医苑(2022年5期)2022-06-10
中国典型病例大全(2022年7期)2022-04-22
科学之谜(2021年2期)2021-04-25
科学导报(2020年54期)2020-09-09
学苑创造·B版(2019年5期)2019-06-14
科学24小时(2019年5期)2019-06-11
女士(2017年3期)2017-03-25
