
地毯草AcMATE1基因的克隆与表达分析
2021-08-26 08:58张郎织李季肤黄杰王志勇陈志坚
热带作物学报 2021年7期
张郎织 李季肤 黄杰 王志勇 陈志坚

摘 要:铝毒被认为是酸性土壤上抑制植物根系生长的重要胁迫因素之一。多药及毒性化合物外排转运蛋白(multidrug and toxic compound extrusion,MATE)在根系柠檬酸转运耐铝毒中起重要作用。以地毯草(Axonopus compressus)为材料,对AcMATE1基因进行克隆、生物信息学和基因表达分析。结果表明:铝处理抑制了地毯草根系的生长,包括总根长、根表面积和根体积。通过PCR克隆到地毯草AcMATE1基因,该基因cDNA全长1602 bp,编码433个氨基酸残基。AcMATE1蛋白分子量为45.9 kDa,属于疏水性蛋白。AcMATE1蛋白包含MatE保守结构域和9个跨膜结构域。亚细胞定位预测分析结果表明,AcMATE1定位于细胞膜上。实时荧光定量PCR结果表明,金属铝(Al)和镉(Cd)处理显著增强AcMATE1基因在地毯草根系中的表达,而镧(La)处理则抑制AcMATE1基因的表达。并且不同铝浓度和时间处理进一步验证Al可增强AcMATE1基因在地毯草根中的表达。此外,AcMATE1基因在地毯草根、茎和叶中均有表达,且在0~1 cm根段中的表达量高于其他根段中的表达量。本研究结果为解析地毯草响应铝毒害的机理提供候选基因资源。
关键词:地毯草;铝毒;MATE转运蛋白;根系;基因表达
中图分类号:Q945.78 文獻标识码:A
Cloning and Expression Analysis of AcMATE1 in Axonopus compressus
ZHANG Langzhi1, LI Jifu1, HUANG Jie1, WANG Zhiyong1*, CHEN Zhijian1,2*
1. College of Forestry, Hainan University / Key Laboratory of Genetics and Germplasm Innovation of Tropical Special Forest Trees and Ornamental Plants, Ministry of Education, Haikou, Hainan 570228, China; 2. Institute of Tropical Crop Genetic Resources, Chinese Academy of Tropical Agriculture Sciences / Key Laboratory of Crop Gene Resources and Germplasm Enhancement in Southern China, Ministry of Agriculture and Rural Affairs, Haikou, Hainan 571101, China
Abstract: Aluminum (Al) toxicity is one of the serious constraints inhibiting root growth in acid soils. The MATE (Multidrug and toxic compound extrusion) transporter plays important roles in citrate exudation for plant Al detoxification. In this study, cloning, bioinformatic and expression analyses of AcMATE1 were performed in carpetgrass (Axonopus compressus). Results showed the that root growth of carpetgrass was inhibited by Al toxicity, as reflected by the decreases in total root length, root surface area and root volume. The full length of AcMATE1 was 1602 bp, encoding 433 amino acid residues. AcMATE1, with protein molecular weight of 45.9 kDa, was predicted to be a hydrophobic protein. Furthermore, AcMATE1 contained one conserved MatE domain and nine transmembrane domains. AcMATE1 was found to be localized on the plasma membrane based on subcellular localization prediction. Quantitative real time PCR analysis showed that the expressions of AcMATE1 were enhanced in the roots of carpetgrass under both Al and Cd treatments, whereas the expression of AcMATE1 was inhibited by La treatment. The transcription level of AcMATE1 increased in the roots of carpetgrass by Al treatments of different concentration and duration. Moreover, the transcripts of AcMATE1 could be detected in roots, stems and leaves, and AcMATE1 exhibited the highest expression in the root tip with 0 to 1 cm in length. This study would provide candidate gene resources for the further dissection of carpetgrass adaptation to Al toxicity.
Keywords: Axonopus compressus; aluminum toxicity; MATE transporter; root; gene expression
DOI: 10.3969/j.issn.1000-2561.2021.07.007
铝(aluminum,Al)是地壳中分布最广泛的金属元素和第3大常见的元素[1]。当土壤pH低于5.0时,铝离子(Al3+)就会从土壤硅酸盐或氧化物中释放出来溶解到土壤溶液中,对植物造成毒害[2]。因此,铝毒被认为是酸性土壤上限制作物生长的主要因素之一。铝离子主要通过破坏细胞核结构、扰乱膜电势和降低细胞壁延展性等方面损害根尖细胞,进而抑制根系生长[3-4]。在长期进化过程中,植物形成了一系列适应铝毒胁迫的机制,如植物根系可通过分泌有机酸、产生粘着液和边缘细胞以及增加根际pH等减少铝进入根尖细胞;进入到细胞中的铝可被有机酸螯合,形成铝-有机酸复合物,或被隔离在液泡中,起到缓解铝毒害的作用[5]。
多药及毒性化合物外排转运蛋白(multidrug and toxic compound extrusion,MATE)属于次级转运蛋白家族[6],通常包含12个跨膜结构域(trans-me-mbrane domain,TMD),少数MATE蛋白含有8~13个TMD[7]。在拟南芥(Arabidopsis thaliana)[8]、葡萄(Vitis vinifera)[9]和煙草(Nicotiana tabacum)[10]等不同植物中相继克隆了MATE转运蛋白基因。研究发现,拟南芥包含56个MATE转运蛋白,分别在铁离子(Fe3+)、类黄酮和柠檬酸等次级代谢产物转运方面发挥作用[11]。如拟南芥AtMATE1作为柠檬酸的转运蛋白参与植物的耐铝过程[12];TaMATE1B在小麦(Triticum aestivum)中具有转运根系柠檬酸缓解铝毒害的功能[13]。
地毯草(Axonopus compressus)属于禾本科地毯草属多年生草本植物,具有植株低矮、易繁殖和成坪性好等优点。地毯草是用于公共绿地、运动场和固土护坡的优良暖季型草坪草[14]。地毯草起源于美国的墨西哥海湾以及热带美洲,在我国,地毯草主要种植于南方热带及亚热带地区[15]。有研究[16]对狗牙根(Cynodon dactylon)、钝叶草(Stenotaohrum secundatum)、假检草(Eremochloa ophiuroides)、结缕草(Zoysia japonica)和地毯草等10种暖季型草坪草进行耐铝性评价分析,发现铝胁迫抑制了大部分草坪草的生长,但地毯草表现出较强的耐铝毒能力。因此,地毯草被认为是挖掘植物适应铝毒胁迫机理的优良禾草植物[17-18]。克隆地毯草根系铝响应基因,对深入解析地毯草耐铝机制具有重要意义。本研究克隆地毯草AcMATE1转运蛋白基因,并对其进行生物信息学和基因表达分析,为解析地毯草适应酸性土壤铝毒机理提供参考。
1 材料与方法
1.1 材料
本研究所用材料为地毯草品系‘A58[19],由海南大学林学院提供。
1.2 方法
1.2.1 材料培养与处理 参考廖丽等[20]的方法进行地毯草移栽和培养。首先将地毯草匍匐茎移植于1/2 Hoagland营养液中预培养5 d,随后,选取生长一致的幼苗(根长约5 cm),在水培箱中进行铝处理。参考廖丽等[20]的方法设置2个铝处理浓度,分别为0 mmol/L和2.1 mmol/L AlCl3,每2 d调节pH至4.2。每个处理设置4个生物学重复,每个重复4株苗。铝处理14 d后收获地毯草,测定根系参数。另一方面,选取生长一致的地毯草幼苗(根长约5 cm),参考Yokosho等[21]的方法在0.5 mmol/L CaCl2液体中进行不同金属处理,包括100 μmol/L AlCl3、40 μmol/L CdCl2和20 μmol/L LaCl3,处理24 h后收获根系用于基因表达分析;选取生长一致的地毯草幼苗(根长约5 cm),在0.5 mmol/L CaCl2液体中进行0、50、100、200 μmol/L AlCl3处理,处理24 h后收获根系用于基因表达分析;在100 μmol/L铝浓度处理下,分别于处理第0、12、24 h收获根系用于基因表达分析。以上处理液pH均调节至4.2,每个处理设置4个生物学重复。
1.2.2 根系参数测定 使用扫描仪(EPSON,日本)扫描并保存根系图像。用根系分析软件WinRHIZO(Regent,加拿大)分析并统计每株总根长、根表面积和根体积。
1.2.3 地毯草总RNA提取和cDNA合成 参照TRIzol(Invitrogen Inc,美国)方法提取铝处理下地毯草根系总RNA。称取0.1 g根系样品,加入1 mL TRIzol提取液,充分捣碎并混匀。加入500 μL氯仿,混匀,静置5 min后,4 ℃,12 000 r/min离心15 min。吸取上清液,加入等体积的异丙醇,充分混匀,4 ℃,12 000 r/min离心10 min。弃上清,加入1 mL 75%乙醇洗涤沉淀,4 ℃,7500 r/min离心5 min。弃上清,吸出残留乙醇,风干沉淀。加入30 ?L无RNase dH2O溶解RNA。参照RevertAid First Strand cDNA Synthesis Kit反转录试剂盒(Thermo Fisher,美国)方法合成cDNA。在PCR管中加入2 μg RNA、1 μL Oligo(dT)18 primer,并加入Water nuclease-free至12 μL。65 ℃反应5 min,冰上冷却。随后加入4 μL 5×Reaction Buffer、1 μL RiboLock RNase Inhibitor、2 μL dTP Mix和1 μL RevertAid M-Mul V Reverse transcriptase,总反应体积为20 μL。反应物置于42 ℃ 60 min,70 ℃ 5 min。反应结束后获得cDNA。
1.2.4 地毯草AcMATE1基因克隆 基于本课题组前期获得的地毯草根系铝响应转录组结果,筛选获得AcMATE1基因全长序列。根据序列设计上下游基因全长开放阅读框(ORF)克隆引物(表1)。以cDNA为模板,通过PCR扩增AcMATE1基因全长。PCR产物经回收、克隆和测序分析后,获得AcMATE1基因全长序列。
1.2.5 生物信息学分析 使用EXPASY(https:// web.expasy.org/protparam)分析蛋白质二级结构、运用DNAMAN软件分析获得氨基酸序列、使用SignalP 5.0 server软件预测信号肽、运用TMHMM 2.0(http://www.cbs.dtu.dk/services/TMHMM)预测跨膜结构域、采用WoLF PSORT(https://www. genscript.com/wolf-psort.html)预测亚细胞定位、利用NCBI(https://www.ncbi.nlm.nih.gov/cdd/)分析保守结构域、采用MEGA 7软件进行系统进化树分析。
1.2.6 实时荧光定量PCR(qPCR)分析 用Quant-StiduoTM Real-Time PCR (Thermo Fisher,美国)和SYBR Green(Vazyme,中国)进行qPCR分析。反应体系包括10 μL 2×SYBR Green PCR master mix、6.4 μL ddH2O、0.8 μL上游引物、0.8 μL下游引物和2 μL模板。定量PCR反应程序为:94 ℃ 1 min;95 ℃ 15 s,58 ℃ 15 s,72 ℃ 30 s,40个循环。基因表达量以AcMATE1基因表达量除以AcEF1a(看家基因)表达量表示。qPCR引物如表1所示。
1.3 数据处理
数据采用Microsoft Excel 2003软件进行计算和制图,采用SPSS软件进行统计分析。
2 结果与分析
2.1 铝处理对地毯草根系生长的影响
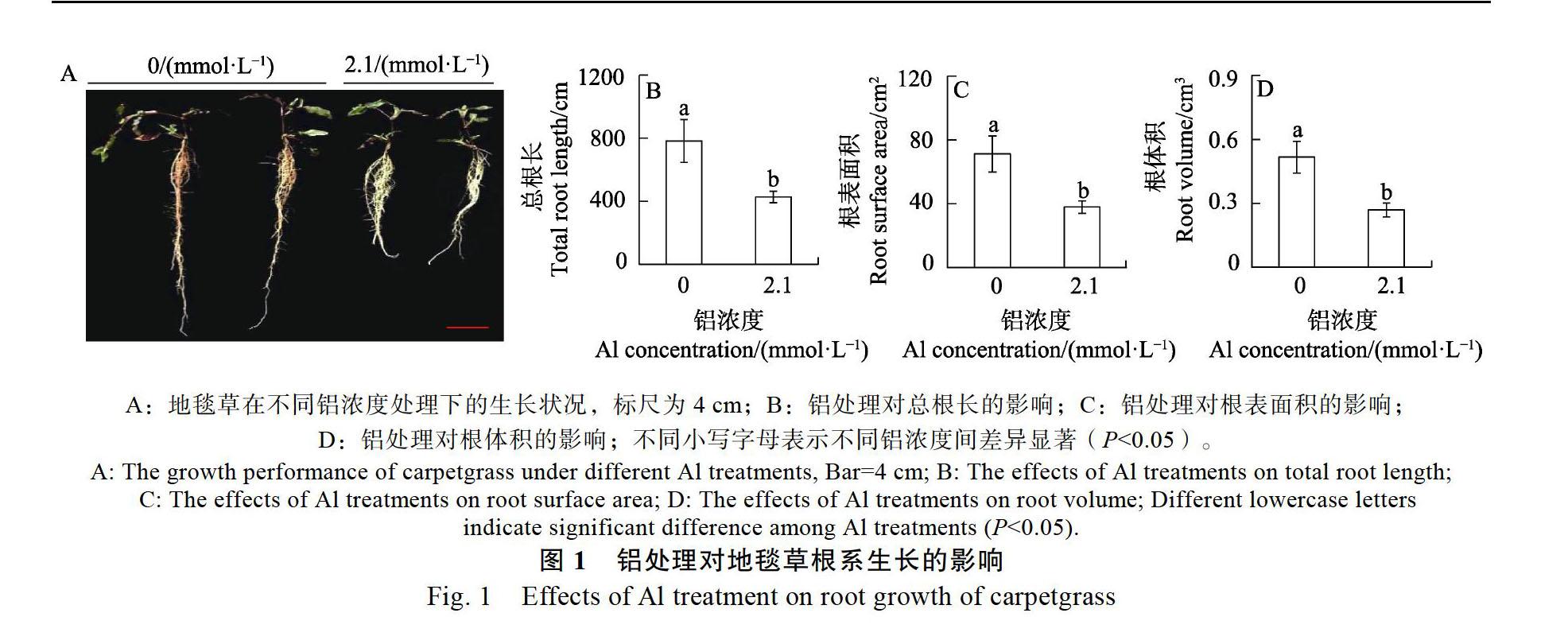
从图1A可看出,2.1 mmol/L铝处理显著抑制地毯草根系生长,2.1 mmol/L铝处理下的地毯草根长明显小于对照(0 mmol/L)根长。2.1 mmol/L铝处理14 d,地毯草总根长、根表面积和根体积分别只有对照处理下的54.6%、53.3%和52.0%(图1B~图1D)。
2.2 AcMATE1基因克隆
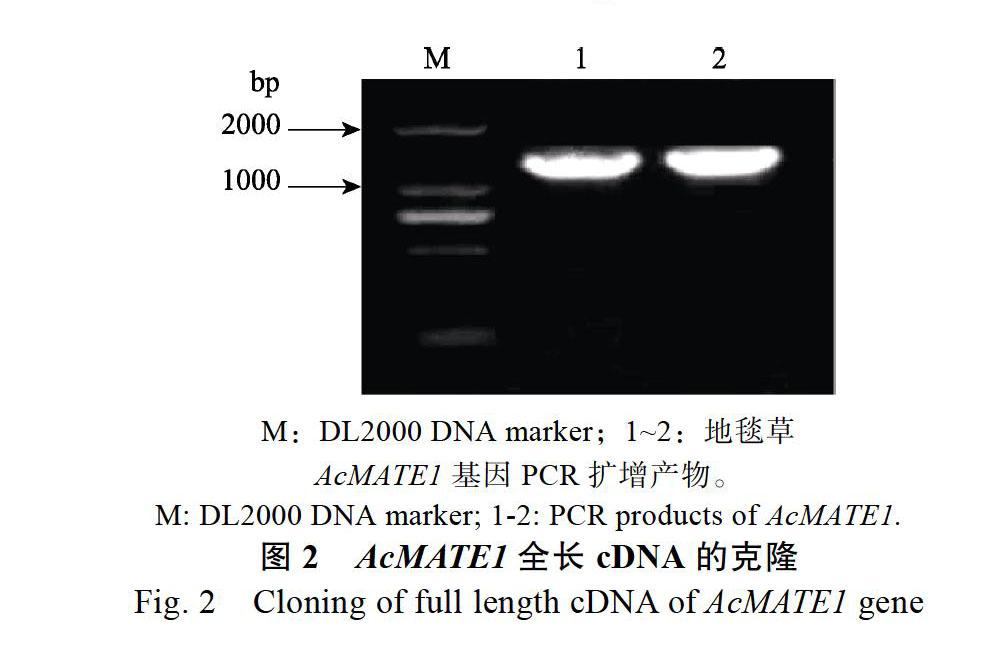
以铝处理下地毯草根系cDNA为模板,PCR扩增出AcMATE1基因的cDNA全长(图2)。PCR产物经克隆测序分析后,获得AcMATE1基因全长核苷酸序列1602 bp。利用DNAMAN和EXPASyProt软件分析获得AcMATE1基因推导的氨基酸序列(图3),该基因编码433个氨基酸残基,蛋白分子量为45.9 kDa,理论等电点为8.3,为疏水性蛋白。
2.3 地毯草AcMATE1蛋白保守结构域分析
CDD Tools预测分析结果表明,AcMATE1属于MATE亚家族成员,包含MATE转运蛋白的保守序列,即MatE结构域和跨膜结构域特征序列(图4)。
2.4 AcMATE1蛋白理化性质分析
运用TMHMM Server V2.0软件进行跨膜结构域分析,发现AcMATE1蛋白包含9個跨膜结构域(图5)。用SignalP 5.0 server软件预测信号肽结构,发现该蛋白不具有信号肽,属于非分泌蛋白。运用WoLF PSORT软件进行亚细胞定位预测,发现该蛋白定位于细胞膜上,暗示该蛋白为膜蛋白。
2.5 AcMATE1同源蛋白系统进化树分析
如图6所示,不同植物MATEs蛋白可被分为Ⅰ、Ⅱ、Ⅲ和Ⅳ共4组。其中,第Ⅰ组包含拟南芥、高粱和水稻等植物的MATE同源蛋白。第Ⅱ组包含拟南芥AtEDS5和AtFRD3、白羽扇豆LaMATE、赤桉EcMATE1和大豆MATE同源蛋白。第Ⅲ组包括烟草NtJAT1、葡萄VvMATE1和拟南芥MATE[22]等同源蛋白。第Ⅳ组包含拟南芥AtTT12、烟草NtMATE1、蒺藜苜蓿和毛果杨MATE同源蛋白。其中,地毯草AcMATE1蛋白被分在第Ⅲ组,与拟南芥AtBCD1和AtADS1等同属1个分支,同源性最高。
2.6 不同处理对AcMATE1基因表达的影响
进一步分析了不同金属元素处理对AcMATE1基因表达的影响,包括100 μmol/L Al、40 μmol/L镉(Cd)和20 μmol/L镧(La)。从图7A可看出,与对照CK处理相比,Al和Cd处理下AcMATE1基因在地毯草根系中的表达均显著增强,且AcMATE1基因在Al处理下的表达量最高,而La处理则抑制了AcMATE1基因的表达。其中,在Al处理下,AcMATE1基因表达量是CK处理下的9.3倍,差异显著(P<0.05)。
如图7B所示,与对照(0 μmol/L Al)相比,随着铝处理浓度的增加,AcMATE1基因表达量呈现增加趋势;在100 μmol/L Al处理下,AcMATE1基因表达量最高,是对照处理下的3.5倍(P< 0.05)。如图7C所示,在100 μmol/L Al处理下,处理时间为12 h和24 h时,AcMATE1基因表达量显著增强,分别是0 h铝处理下的23.1倍和12.2倍(P<0.05)。
2.7 AcMATE1基因组织表达分析
组织表达模式分析结果发现(图8A),地毯草AcMATE1基因在根、茎和叶中均有表达,其中, AcMATE1基因在叶中的表达量最高,根和茎次之。另外,我们分析了AcMATE1基因在不同根段中的表达,结果发现AcMATE1基因在0~1 cm根段中的表达量显著高于在1~2 cm和3 cm以上根段中的表达量(图8B)。
3 討论
铝毒是限制作物生长和产量的重要因素之一,通过阻碍细胞分裂和伸长而抑制根系生长[23]。不同植物或同一植物不同品种对铝毒害忍耐能力存在差异。本研究以地毯草为材料,首先分析了铝处理对地毯草根系生长的影响。结果发现铝处理抑制了地毯草的总根长、根表面积和根体积。在其他植物中也发现铝处理显著抑制根系的生长。例如,在拟南芥中,5 μmol/L铝处理能够明显抑制主根长的伸长[24]。在大麦不同基因型中,铝处理均能抑制根系生长[25]。另外,水稻根系的生长也受到铝毒害的影响[[21]。虽然铝处理抑制了不同草坪草的生长,但相对于钝叶草、结缕草和假检草等,地毯草具有较强的耐铝能力[16]。因此,为更好地探讨地毯草适应酸性土壤铝毒害机制,本研究进一步克隆了地毯草AcMATE1基因,并对其进行生物信息学及基因表达分析。
地毯草AcMATE1蛋白属于MATE转运蛋白家族成员,具有高度保守的MatE结构域[7]。近年来,已经克隆并鉴定出多种植物的MATE蛋白。例如,在拟南芥中,包含有56个AtMATEs[26],在水稻中,有53个OsMATEs[27],而蒺藜苜蓿(Medicago truncatula)和大豆(Glycine max)中分别包含有70和117个MATEs[28-29]。本研究发现,地毯草AcMATE1蛋白具有MatE保守结构域,且含有9个TMD(图5)。植物MATE蛋白主要定位于质膜或液泡膜上,这对其行使生物学功能至关重要[26]。亚细胞定位预测发现,地毯草AcMATE1定位于质膜上,属于膜蛋白。以上结果表明,AcMATE1具备植物MATE蛋白的典型特征。
植物MATE蛋白在激素信号转递和金属离子转运中起重要作用。例如,拟南芥AtADS1是一类MATE转运蛋白,超量表达AtADS1能降低拟南芥水杨酸水平,进而影响了拟南芥的抗病能力[29]。拟南芥高尔基体定位的MATE蛋白(AtBCD1),具有通过调节铁的分配来维持胁迫条件下细胞内铁平衡的生物学功能[30]。在本研究中,地毯草AcMATE1蛋白与拟南芥AtBCD1和AtADS1具有较高的同源性,暗示了地毯草AcMATE1蛋白可能通过调控植物激素和金属离子转运响应铝毒胁迫。此外,植物MATE蛋白在柠檬酸转运中也具有重要作用。例如,赤小豆VuMATE1蛋白定位于质膜上,其被报道具有调控根尖分泌柠檬酸的生物学功能[31];水稻OsFRDL1具有转运柠檬酸的功能,同时也调控了铁在水稻谷粒中的分配[32]。
本研究进一步发现铝处理显著增强了AcMATE1基因在地毯草根系中的表达,且AcMATE1基因在0~1 cm根段中表达量最高。ZmMATE1基因具有转运柠檬酸的功能,铝处理诱导了ZmMATE1在玉米根中的表达,且ZmMATE1基因在耐铝玉米基因型中的表达量高于铝敏感基因型[33]。此外,铝处理也增强了BoMATE在甘蓝根中的表达,超量表达BoMATE能促进转基因拟南芥根系分泌柠檬酸,从而增强拟南芥的耐铝能力[34]。研究发现,铝处理增强了OsFRDL4基因在水稻根尖中的表达,且OsFRDL4基因表达与水稻耐铝能力呈正相关关系;敲除OsFRDL4基因显著降低了水稻根系柠檬酸的分泌量,并降低了水稻的耐铝能力,表明OsFRDL4是水稻耐铝毒的重要基因[21]。然而,本研究发现AcMATE1基因在地毯草叶中的表达量高于根部,暗示其可能主要在叶部中发挥功能,而铝处理增强了AcMATE1基因在地毯草根中的表达,暗示了该基因也能够响应铝毒害,其部分功能可能参与了地毯草根系响应铝毒害的过程,但其具体的生物学功能仍需要做进一步研究。本研究为探讨地毯草耐铝毒机理提供了候选基因资源。
参考文献
[1] Teng W C, Kang Y C, Hou W J, et al. Phosphorus application reduces aluminum toxicity in two Eucalyptus clones by increasing its accumulation in roots and decreasing its content in leaves[J]. PLoS One, 2018, 13(1): e0190900.
[2] Pereira J F, Ryan P R. The role of transposable elements in the evolution of aluminium resistance in plants[J]. Journal of Experimental Botany, 2019, 70(1): 41-54.
[3] Ma J F, Chen Z C, Shen R F. Molecular mechanisms of Al tolerance in gramineous plants[J]. Plant and Soil, 2014, 381(1-2): 1-12.
[4] Kochian L V, Pi?eros M A, Liu J, et al. Plant adaptation to acid soils: the molecular basis for crop aluminum resis?ta-nce[J]. Annual Review of Plant Biology, 2015, 66: 571-598.
[5] Sun L L, Zhang M S, Liu X M, et al. Aluminium is essential for root growth and development of tea plants (Camellia sinensis)[J]. Journal of Integrative Plant Biology, 2020, 62(7): 984-997.
[6] Takanashi K, Shitan N, Yazaki K, et al. The multidrug and toxic compound extrusion (MATE) family in plants[J]. Plant Biotechnology, 2014, 31(5): 417-430.
[7] Magalhaes J V. How a microbial drug transporter became essential for crop cultivation on acid soils: aluminium tolera?n?ce conferred by the multidrug and toxic compound extrusion (MATE) family[J]. Annals of Botany, 2010, 106(1): 199-203.
[8] Roschzttardtz H, Séguéla-Arnaud M, Briat J F, et al. The FRD3 citrate effluxer promotes iron nutrition between symplastically disconnected tissues throughout Arabidopsis develo?pment[J]. Plant Cell, 2011, 23(7): 2725-2737.
[9] Pérez-Díaz R, Ryngajllo M, Pérez-Díaz J, et al. VvMATE1 and VvMATE2 encode putative proanthocyanidin transporters expressed during berry development in Vitis vinifera L.[J]. Plant Cell Reports, 2014, 33(7): 1147-1159.
[10] Shitan N, Minami S, Morita M, et al. Involvement of the leaf- specific multidrug and toxic compound extrusion (MATE) transporter Nt-JAT2 in vacuolar sequestration of nicotine in Nicotiana tabacum[J]. PLoS One, 2014, 9(9): e108789.
[11] Thompson E P, Wilkins C, Demidchik V, et al. An Arabidopsis flavonoid transporter is required for anther dehiscence and pollen development[J]. Journal of Experimental Botany, 2010, 61(2): 439-451.
[12] Liu J, Luo X, Shaff J, et al. A promoter-swap strategy between the AtALMT and AtMATE genes increased Arabid-opsis aluminum resistance and improved carbon-use efficiency for aluminum resistance[J]. The Plant Journal, 2012, 71(2): 327-337.
[13] Tovkach A, Ryan P R, Richardson A E, et al. Transposon-mediated alteration of TaMATE1B expression in wheat confers constitutive citrate efflux from root apices[J]. Plant Physiology, 2013, 161(2): 880-892.
[14] 廖 丽, 陈玉華, 赵亚荣, 等. 地毯草种质资源形态多样性[J]. 草业科学, 2015, 32(2): 248-257.
[15] 李洁英, 解安霞, 白昌军, 等. 周期性去叶对地毯草克隆生长的影响[J]. 草业学报, 2011, 20(3): 115-121.
[16] Baldwin C M, Liu H, Mccarty L B, et al. Aluminum tolerances of ten warm-season turfgrasses[J]. Golfdom, 2005, 10: 881-887.
[17] 张 静, 廖 丽, 白昌军, 等. 地毯草耐铝性初步评价[J]. 草业科学, 2012, 29(11): 1671-1677.
[18] 李季肤, 韩佳芮, 贾怡丹, 等. 地毯草铝响应基因AcABCG1的克隆与表达分析[J]. 草地学报, 2019, 27(5): 1147-1153.
[19] 劉 洋, 廖 丽, 阙 勇, 等. 地毯草种质资源耐铝性评价[J]. 热带作物学报, 2017, 38(4): 602-610.
[20] 廖 丽, 黄小辉, 白昌军, 等. 地毯草对铝胁迫响应及临界浓度的研究[J]. 热带作物学报, 2011, 32(7): 1235-1239.
[21] Yokosho K, Yamaji N, Ma J F. An Al-inducible MATE gene is involved in external detoxification of Al in rice[J]. The Plant Journal, 2011, 68(6): 1061-1069.
[22] Liu J, Li Y, Wang W, et al. Genome-wide analysis of MATE transporters and expression patterns of a subgroup of MATE genes in response to aluminum toxicity in soybean[J]. BMC Genomics, 2016, 17: 223.
[23] Famoso A N, Clark R T, Shaff J E, et al. Development of a novel aluminum tolerance phenotyping platform used for comparisons of cereal aluminum tolerance and investigations into rice aluminum tolerance mechanisms[J]. Plant Physio?logy, 2010, 153(4): 1678-1691.
[24] Yang Z B, Liu G C, Liu J J, et al. Synergistic action of auxin and cytokinin mediates aluminum-induced root growth inhibition in Arabidopsis[J]. EMBO Reports, 2017, 18(7): 1213-1230.
[25] Ahmed I M, Nadira U A, Cao F B, et al. Physiological and molecular analysis on root growth associated with the tolera?n?ce to aluminum and drought individual and combined in Tibetan wild and cultivated barley[J]. Planta, 2016, 243(4): 973-985.
[26] Wang L H, Bei X J, Gao J S, et al. The similar and different evolutionary trends of MATE family occurred between rice and Arabidopsis thaliana[J]. BMC Plant Biology, 2016, 16(1): 207.
[27] Tiwari M, Sharma D, Singh M, et al. Expression of OsMATE1 and OsMATE2 alters development, stress respo-nses and pathogen susceptibility in Arabidopsis[J]. Scientific Reports, 2014, 4: 3964.
[28] Wang J J, Hou Q Q, Li P H, et al. Diverse functions of multid?rug and toxin extrusion (MATE) transporters in citric acid efflux and metal homeostasis in Medicago truncatula[J]. The Plant Journal, 2017, 90(1): 79-95.
[29] Sun X L, Gilroy E M, Chini A, et al. ADS1 encodes a MATE-transporter that negatively regulates plant disease resistance[J]. New Phytologist, 2011, 192(2): 471-482.
[30] Seo P J, Park J, Park M J, et al. A Golgi-localized MATE tra?nsporter mediates iron homoeostasis under osmotic stress in Arabidopsis[J]. Biochemical Journal, 2012, 442(3): 551- 561.
[31] Yang X Y, Yang J L, Zhou Y, et al. A de novo synthesis citrate transporter, Vigna umbellata multidrug and toxic compound extrusion, implicates in Al-activated citrate efflux in rice bean (Vigna umbellata) root apex[J]. Plant Cell and Environment, 2011, 34(12): 2138-2148.
[32] Yokosho K, Yamaji N, Ma J F. OsFRDL1 expressed in nodes is required for distribution of iron to grains in rice[J]. Journal of Experimental Botany, 2016, 67(18): 5485-5494.
[33] Maron L G, Pi?eros M A, Guimar?es C T, et al. Two functi?o?nally distinct members of the MATE (multi-drug and toxic compound extrusion) family of transporters potentially und?erlie two major aluminum tolerance QTLs in maize[J]. The Plant Journal, 2010, 61(5): 728-740.
[34] Wu X X, Li R, Shi J, et al. Brassica oleracea MATE encodes a citrate transporter and enhances aluminum tolerance in Arabidopsis thaliana[J]. Plant and Cell Physiology, 2014, 55(8): 1426-1436.
責任编辑:黄东杰
猜你喜欢
廉政瞭望·下半月(2022年4期)2022-05-12
汉语世界(The World of Chinese)(2019年2期)2019-04-19
小火炬·智漫悦读(2017年7期)2017-09-29
中文信息(2017年2期)2017-04-13
江苏农业科学(2016年11期)2017-03-21
足球周刊(2016年15期)2016-11-02
智能计算机与应用(2016年4期)2016-09-26
飞碟探索(2015年9期)2015-11-05
红领巾·探索(2015年9期)2015-09-10
恋爱婚姻家庭·养生版(2011年8期)2011-05-14
