
油茶果生刺盘孢液泡分选蛋白CfVps26的功能*
2021-10-09 05:49李茜雅张盛培
林业科学 2021年8期
李茜雅 张盛培 李 河
(经济林培育与保护省部共建教育部重点实验室 中南林业科技大学 长沙 410004)
油茶(Camelliaoleifera)树不仅可以保持水土、涵养水源,而且还能提供优质的食用油,具有极高的生态、经济和社会价值(刘跃进等, 2007; 陈永忠等, 2013)。由刺盘孢属(Colletotrichumspp.)真菌导致的炭疽病能造成油茶落叶、落花和落果(靳爱仙等, 2009)。油茶籽产量因炭疽病减产10%~30%,重病区甚至减产50%以上,造成巨大的经济损失(徐丽萍等, 2015)。目前,我国防治油茶炭疽病主要采用苯并咪唑类(多菌灵或甲基托布津等)内吸性杀菌剂(郑少华, 2012),但由于其作用位点和机制较为单一,容易使病原菌产生抗药性。而病原菌对多靶标位点的杀菌剂不易产生抗药性(詹家绥等, 2014),因此亟需研究筛选出更多新的药物靶标位点,为新型杀菌剂的研发提供依据。
Retromer复合体主要介导跨膜蛋白从内涵体到反式高尔基体的运输和从内涵体到细胞膜的运输,使得蛋白受体可以反复循环利用,避免运输到溶酶体被降解,维持细胞内的代谢平衡(Bonifacinoetal., 2008; Hierroetal., 2007)。该复合体由液泡分选蛋白(vacuolar protein sorting,简称vps)组成,分为由Vps26、Vps29和 Vps35构成的货物识别三聚体和由Vps5和Vps17构成的微管分选蛋白二聚体(Cullenetal., 2011)。禾谷镰刀菌(Fusariumgraminearum)Retromer复合体Vps5、Vps17、Vps26、Vps29和 Vps35五个分选蛋白均参与调控菌丝生长、产孢和致病过程(张莹, 2011)。在稻瘟病菌(Magnaporthegrisea)Vps26、Vps29和 Vps35组成的货物识别三聚体均影响该病菌的产孢量和致病力(郑文辉, 2014)。
果生刺盘孢(Colletotrichumfructicola)是油茶炭疽病优势致病菌(李河等, 2014; 2017; 2018; 2019)。笔者课题组从果生刺盘孢全基因组中鉴定到1个与酿酒酵母(Saccharomycescerevisiae)的Retromer复合体亚基Vps26的同源蛋白,命名为CfVps26,但其生物学功能尚不清楚。本文拟以液泡分选蛋白Vps26为研究对象,其在油茶果生刺盘孢中的生物学功能,为探究Vps26的作用机制和防治油茶炭疽病提供参考。
1 材料与方法
1.1 供试菌株
果生刺盘孢CFLH16为野生型菌株,突变体ΔCfvps26和回补菌株ΔCfvps26/CfVPS26由本试验获得。
1.2 CfVps26蛋白系统发育分析
根据酿酒酵母中Vps26蛋白氨基酸序列(XP_031879784.1)在果生刺盘孢全基因组数据库(ANPB00000000.1)中进行搜索,获得与其同源的CfVps26蛋白氨基酸序列,与NCBI数据库中其他真菌的Vps26蛋白氨基酸序列进行系统发育分析。
1.3 CfVPS26基因敲除载体构建及突变体筛选
以野生型菌株 CFLH16 基因组 DNA 为模板,设计引物CfVPS26-1F/CfVPS26-2R、CfVPS26-3F/CfVPS26-4R(表1)分别PCR扩增CfVPS26目的基因上下游各1 kb左右的DNA片段,将其作为构建敲除载体片段的上下臂。引物 Hyg F/Hyg R(表1)用来扩增潮霉素抗性基因片段(HPH)。利用引物CfVPS26-1F/CfVPS26-4R,采用Over-lap方法将上臂、潮霉素和下臂DNA片段顺序连接,得到CfVPS26基因敲除载体片段。
参照Zhang等(2019)方法制备果生刺盘孢CFLH16原生质体。参考高亚兰等(2020)的方法将CfVPS26基因敲除载体片段转化至原生质体中。采用基因内探针引物CfVPS26-7F/CfVPS26-8R和基因外引物CfVPS26-5F/H855R(表1)验证转化子。基因内探针引物不能扩增出目的基因条带,且基因外引物可扩增出正确大小条带的转化子为CfVPS26基因敲除突变体。

表1 CfVPS26在果生刺盘孢研究中使用的引物Tab.1 CfVPS26 primers used in Colletotrichum fructicola study
1.4 CfVPS26基因敲除突变体回补菌株的获得
采用引物CfVPS26-9F/CfVPS26-10R PCR扩增含有启动子在内的CfVPS26基因回补片段,参照高亚兰等(2020)方法构建回补质粒; 将回补质粒转化至突变体ΔCfvps26原生质体中,在添加博来霉素的TB3培养基上培养3 天,荧光筛选回补菌株。
1.5 突变体ΔCfvps26的表型测定
1.5.1 不同种类培养基上生长测定 参照姚权等(2019)方法对菌株的生长速率进行测定。
1.5.2 细胞壁敏感性测定 在野生型、突变体和回补菌株菌落边缘用无菌打孔器(Φ=8 mm)切取菌饼,分别接种含0.01%十二烷基硫酸钠(sodium dodecyl sulfate,SDS)和200 μg·mL-1荧光增白剂(calcofluor white,CFW)的PDA平板上,培养3天后测量菌落直径。
1.5.3 产孢量与附着胞形成测定 将野生型、突变体和回补菌株菌落分别接种在PDB培养基中,培养4 天后统计产孢量。得到等浓度(105个·mL-1)分生孢子悬浮液,滴加10 μL 置于疏水玻片上,保湿培养12 h后观察记录附着胞形成率情况。
1.5.4 致病力测定 在培养3天的野生型、突变体和回补菌株菌落边缘用无菌打孔器(Φ=8 mm)切取菌饼,将菌饼有菌丝的一面接种油茶叶片上,空白PDA培养基作为对照。保湿培养,观察发病情况并统计直径。
1.5.5 内质网压力胁迫剂敏感性测定 在野生型、突变体和回补菌株菌落边缘用经灭菌的打孔器(Φ=8 mm)切取菌饼,接种于含5 mmol·L-1二硫苏糖醇DTT的PDA平板上,培养4 天,测量菌落直径。
1.5.6 糖原染色 参照郑文辉(2014)的方法对附着胞进行糖原染色,制备浓度为105个·mL-1的野生型CFLH16和突变体ΔCfvps26分生孢子悬浮液,各取10 μL滴于疏水玻片上黑暗保湿培养24 h,吸取多余水分后滴加30 μL糖原染料(I2/KI,120 mg·mL-1KI + 20 mg·mL-1I2),黑暗静止染色30 min,显微镜观察附着胞糖原代谢情况。
2 结果与分析
2.1 CfVps26的鉴定及系统进化分析
鉴定到1个与酿酒酵母中Retromer复合体亚基Vps26同源的蛋白CfVps26,对其结构域进行分析发现,从第5~279个氨基酸仅有1个Vps26结构域(图1A)。系统发育分析表明果生刺盘孢中Vps26氨基酸序列与希金斯刺盘孢(C.higginsianum)亲缘关系最近,与酿酒酵母亲缘关系最远(图1B)。
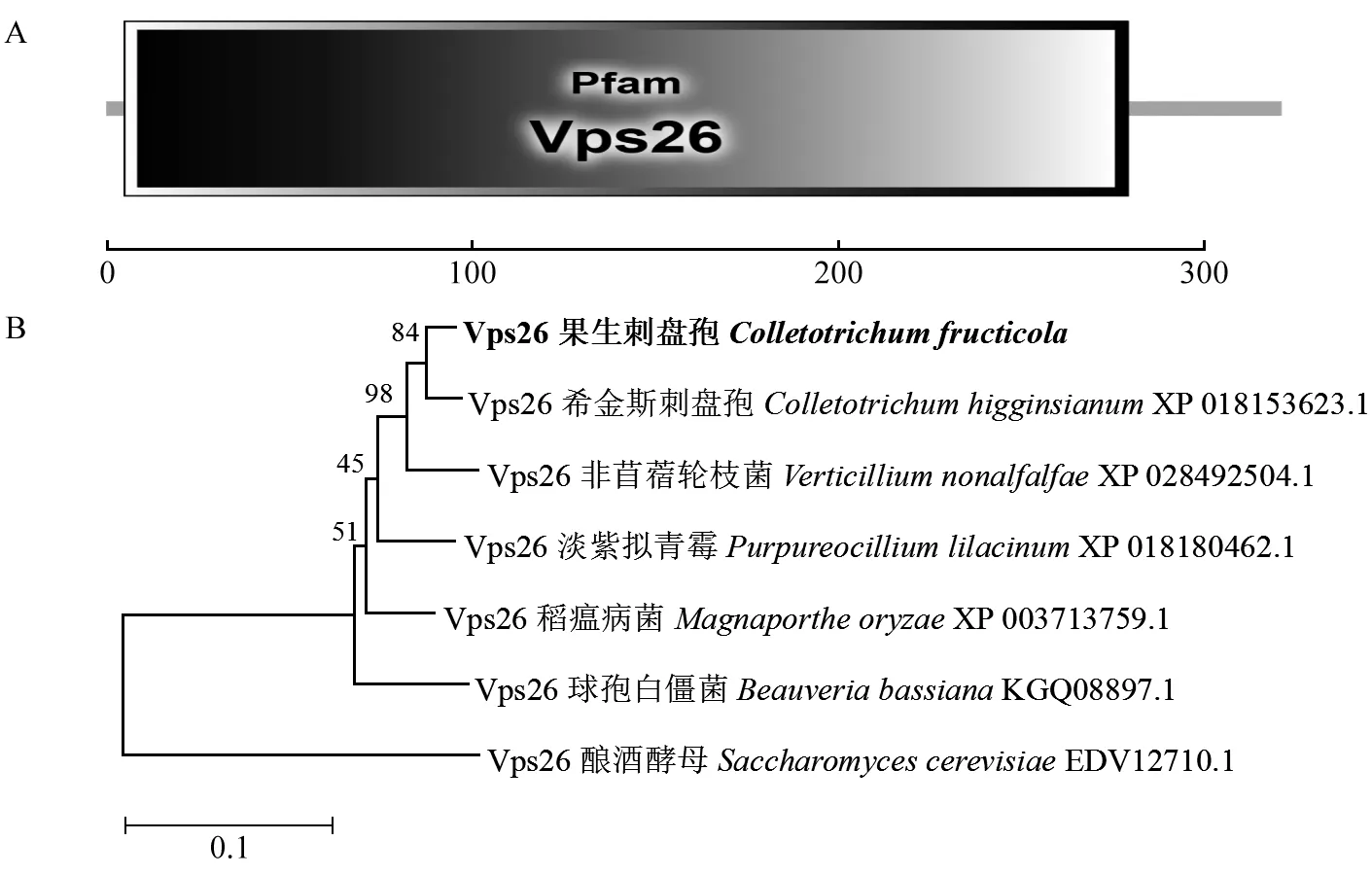
图1 CfVps26结构域和系统发育分析Fig. 1 Domain and phylogenetic analysis of CfVps26A: CfVps26蛋白的结构域。B: 邻近相连法构建系统发育树。A: The structure of CfVps26; B: Phylogenetic tree was constructed by neighbor joining method.
2.2 CfVPS26基因敲除突变体及回补菌株的获得
根据同源重组原理构建CfVPS26基因敲除策略(图2A)。利用引物CfVPS26-7F/CfVPS26-8R和CfVPS26-5F/H855R进行验证,获得突变体ΔCfvps26。回补菌株ΔCfvps26/CfVPS26通过荧光筛选以及PCR验证获得(图2B)。

图2 获得CfVPS26基因敲除突变体Fig. 2 Generation of the CfVPS26 gene deletion mutantA: 基因敲除策略图; B: 电泳验证图。A: Schematic of the deletion strategy; B: Electrophoregram for verification.
2.3 CfVPS26参与调控果生刺盘孢营养生长、产孢和附着胞形成
突变体ΔCfvps26在PDA和MM培养基上,菌落直径均小于野生型及回补菌株(图3A),统计分析差异显著(图3B),气生菌丝明显减少(图3C),说明CfVPS26能够调控果生刺盘孢菌体的营养生长。

图3 CfVPS26基因敲除突变体的菌丝生长测定和产孢量、附着胞形成率统计分析Fig. 3 Growth defect, statistical analysis of conidial quantity and appressorium formation rate of CfVPS26 gene deletion mutantA: 突变体在PDA、MM 培养基上生长情况; B: 直径统计; C: 菌落侧视图; D: 突变体ΔCfvps26能形成少量附着胞; E:产孢量统计; F:附着胞形成率。 不同小写字母表示不同菌株间差异极显著(P≤0.01)。下同。A: Growth of ΔCfvps26 strain inoculated on PDA and MM media; B: Diameter; C: Lateral views; D: The ΔCfvps26 can form a small amount appressorium; E: Conidiation; F: Appressorium formation rate. Different small letters indicate significan difference between different strains(P≤0.01). The same below.
对野生型菌株CFLH16、突变体ΔCfvps26以及回补菌株ΔCfvps26/CfVPS26的产孢量进行统计分析,发现突变体菌株ΔCfvps26产孢量显著下降,只有野生型和回补菌株产孢量的10%左右(图3E)。附着胞是病原真菌侵染植物组织的关键结构。对野生型菌株CFLH16、突变体ΔCfvps26以及回补菌株ΔCfvps26/CfVPS26的附着胞形成率统计分析发现,与野生型和回补菌株相比,突变体ΔCfvps26附着胞形成率显著下降(图3D),且差异显著(图3F)。以上结果说明CfVPS26基因参与调控果生刺盘孢分生孢子及附着胞的形成,这可能是突变体ΔCfvps26对油茶叶片致病力显著降低的主要原因。
2.4 CfVPS26参与调控果生刺盘孢对外界环境胁迫的应答
真菌在生长发育和致病过程中会受到多种外界环境胁迫的影响,进一步研究CfVPS26基因是否响应细胞壁胁迫应答,将野生型菌株CFLH16、突变体ΔCfvps26和回补菌株ΔCfvps26/CfVPS26分别接种于含有细胞壁抑制剂CFW和SDS的 PDA培养基上,比较生长情况。结果表明,突变体ΔCfvps26对CFW和SDS敏感性增强,其生长抑制率高于野生型和回补菌株,且差异显著(图4A、B),说明CfVPS26基因参与果生刺盘孢对细胞壁胁迫应答。
为确定CfVPS26基因是否参与调控内质网压力胁迫应答过程,将野生型菌株CFLH16、突变体菌株ΔCfvps26和回补菌株ΔCfvps26/CfVPS26分别接种于含有5 mmol·L-1二硫苏糖醇DTT的PDA培养基上,比较生长抑制率。结果表明,突变体菌株ΔCfvps26对DTT耐受性显著高于野生型菌株和回补菌株(图4C、D),说明CfVPS26基因负调控果生刺盘孢应答内质网压力胁迫过程。

图4 突变体ΔCfvps26参与外界环境胁迫应答Fig. 4 Growth assays of the ΔCfvps26 mutant on PDA with environmental stressorsA: 突变体ΔCfvps26在细胞壁胁迫下的生长情况; B: 抑制率统计; C:突变体ΔCfvps26在DTT胁迫下的生长情况; D: 抑制率统计。A: Growth of ΔCfvps26 under cell wall stress; B: Inhibition rates; C: Growth of ΔCfvps26 under DTT stress; D: Inhibition rates.
2.5 CfVPS26基因参与调控果生刺盘孢的致病力
对突变体ΔCfvps26、野生型和回补菌株进行了致病力测定,结果显示,在离体接种3天后,与野生型和回补菌株相比突变体ΔCfvps26没有产生病斑,丧失了对油茶叶片的致病力(图5A、B)。以上结果说明CfVPS26基因参与调控果生刺盘孢的致病力。
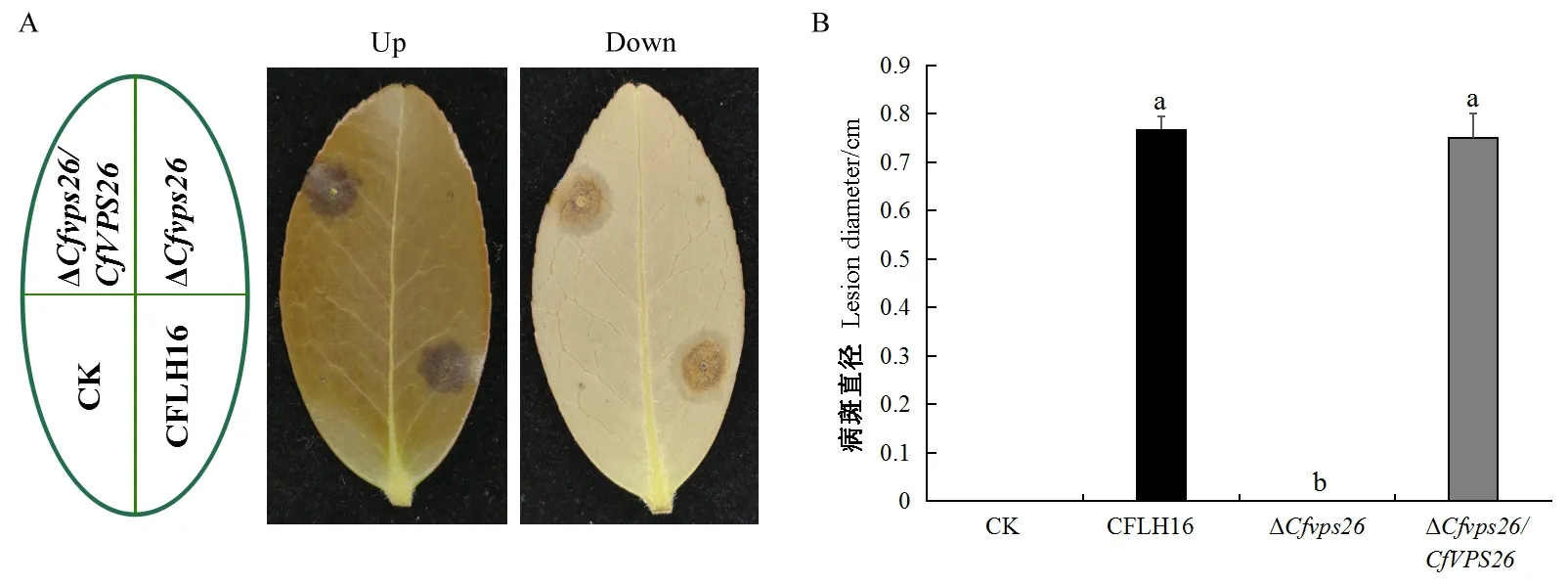
图5 突变体ΔCfvps26对油茶叶片的致病力测定Fig. 5 Pathogenicity of ΔCfvps26 to oil-tea leavesA: 野生型、突变体ΔCfvps26和回补菌株接种在有伤叶片上; B: 病斑统计。A: CFLH16, ΔCfvps26 and ΔCfvps26/CfVPS26 were inoculated onto wounded leaves; B: Lesion diameter.
2.6 CfVPS26基因参与调控果生刺盘孢糖原代谢
附着胞内膨压的形成来源于甘油的积累。分生孢子的萌发后,糖原物质会逐渐从分生孢子向附着胞转移,随后发生降解进而转变为甘油(Wangetal., 2005)。本试验利用糖原染色剂I2/KI对分生孢子及附着胞进行染色,结果显示,野生型菌株分生孢子和附着胞糖原已经几乎代谢完全,而突变体ΔCfvps26中仍然有大量的糖原存在(图6),说明突变体中糖原代谢受阻。结果表明,CfVPS26基因参与调控果生刺盘孢的糖原代谢过程。

图6 糖原染色Fig. 6 Staining for glycogen
3 讨论
3.1 CfVPS26基因与营养生长的关系
菌丝是丝状真菌营养生长的主要模式,侵染菌丝可以在寄主组织内扩展定殖。VPS26基因参与调控植物病原真菌菌丝生长,张莹(2011)研究发现禾谷镰刀菌(Fusariumgraminearum)中VPS26基因敲除突变体生长速率减慢、气生菌丝明显减少。本研究发现,果生刺盘孢的CfVPS26基因敲除后,突变体ΔCfvps26菌丝生长速率减慢,气生菌丝量也显著少于野生型和回补菌株。但黄清平(2013)研究发现,稻瘟病菌(Magnaportheoryzae)中的VPS26基因敲除后,其菌丝形态和生长速率则无明显变化。因此VPS26基因在调控不同病原菌的菌丝生长时作用并不完全相同。
3.2 CfVPS26基因与内质网压力的关系
内质网是蛋白质合成和分泌的重要场所,其稳态平衡在细胞生长发育中具有重要作用。当内质网负担过重或压力过大时,会导致蛋白质错误折叠,此时,细胞便会通过非折叠蛋白反应(unfolded protein response, UPR)来恢复内质网的正常功能。汤蔚(2015)研究发现,稻瘟菌的bZIP 转录因子Hac1和跨膜蛋白激酶IRE1均参与调控病菌内质网应激过程。笔者实验室前期研究也表明,果生刺盘孢CfHAC1基因敲除突变体对5 mmol·L-1二硫苏糖醇耐受性下降,说明转录因子CfHac1正调控病菌对内质网压力胁迫应答过程(李司政等, 2020)。而本研究中,笔者发现果生刺盘孢的CfVPS26基因敲除突变体ΔCfvps26对5 mmol·L-1二硫苏糖醇耐受性增强,说明该基因在应答二硫苏糖醇造成的内质网压力过程中发挥负调控作用,但其调控分子机制有待进一步深入研究。
3.3 CfVPS26基因与分生孢子产生的关系
植物病原真菌的分生孢子作为初侵染与再侵染的接种体,借气流传播从寄主表皮直接侵入致病。VPS26基因参与了植物病原真菌分生孢子的形成,对稻瘟病菌中基因VPS26进行敲除后,突变体菌株分生孢子产量显著下降(黄清平, 2013)。而禾谷镰刀菌中VPS26基因敲除突变体则几乎不产孢或产生畸形孢子(张莹, 2011)。在本研究中,笔者发现果生刺盘孢的突变体菌株ΔCfvps26的分生孢子产量也显著下降。另外,笔者实验室前期研究发现,果生刺盘孢中CfPMK1、CfHAC1、CfMKK1等基因均参与调控分生孢子的形成过程(李河, 2018; 姚权等, 2019; 肖宇等, 2021),说明在果生刺盘孢中存在多种基因参与调控分生孢子的形成这一过程。
3.4 CfVPS26基因与附着胞发育的关系
附着胞是侵染寄主的关键结构(Bechingeretal.,1999)。Kitade等(2015)研究发现玉米(Zeamays)小斑病菌(Bipolarismaydis)的STE7基因参与调控附着胞的形成。在胶胞刺盘胞(C.gloeosporioides)中的CgRGS7基因也影响了附着胞的形成过程(吴曼莉等, 2017)。前期对果生刺盘孢的研究发现,CfPMK1、CfSNF1和CfMKK1基因缺失突变体都不能形成附着胞(李河, 2018; Zhangetal., 2019; 肖宇等, 2021)。在本研究发现,突变体ΔCfvps26附着胞形成率显著降低,这可能是导致其致病力丧失的原因之一。然而黄清平(2013)的研究发现稻瘟病菌中突变体ΔMoVps26对分生孢子的萌发和附着胞形成并无决定性作用,说明VPS26基因在不同的病原菌种类中功能存在差异。
3.5 附着胞发育与糖原代谢的关系
附着胞正常的膨压对病原菌侵入寄主十分重要(Jongetal., 1997)。膨压的产生需要大量甘油的累积,它来源于分生孢子内的糖原等能源物质,通过芽管运输至附着胞降解形成甘油(Wangetal., 2005; Talbot, 2003)。在希金斯刺盘孢中,ChODC基因参与调控附着胞中糖原的转移过程(严亚琴, 2020)。稻瘟病菌中基因MoEND3和MoCRN的缺失会导致糖原转移和降解非常缓慢(李潇, 2017)。在本研究中,果生刺盘孢野生型菌株CFLH16的糖原随着附着胞发育成熟而逐渐代谢,而突变体ΔCfvps26分生孢子中仍有糖原滞留,附着胞内糖原代谢显著减慢,说明糖原在突变体ΔCfvps26中的代谢受到了限制。由此推测CfPVS26基因参与了糖原的代谢过程,其敲除突变体不能形成正常的附着胞膨压,进而丧失了对油茶叶片的致病力。
4 结论
本研究在油茶果生刺盘孢中鉴定到1个Retromer逆向囊泡转运复合体亚基蛋白CfVps26,研究发现该蛋白参与调控病菌生长发育、产孢、附着胞形成、糖原代谢、外界胁迫应答和致病过程,研究结果有助于揭示果生刺盘孢致病的分子机制,并为开发杀菌剂提供潜在药靶。
猜你喜欢
广西糖业(2022年5期)2022-11-24
亚热带农业研究(2022年1期)2022-08-08
蔬菜(2021年7期)2021-11-27
疯狂英语·新策略(2021年9期)2021-11-02
中国农业科技导报(2020年3期)2020-03-15
山西农业科学(2020年2期)2020-02-29
热带作物学报(2019年4期)2019-06-11
安徽农学通报(2017年23期)2017-12-27
成都体育学院学报(2017年1期)2017-02-21
求医问药(2009年7期)2009-08-31
