
橘小实蝇雌雄成虫生殖系统中可培养细菌的分离鉴定
2021-10-20 13:28杨海燕胡凯平宋水林王广利王建国李小珍
环境昆虫学报 2021年5期
杨海燕,胡凯平,宋水林,王广利,徐 业,王建国,李小珍*
(1.江西农业大学农学院,南昌 330045;2.江西农业大学产业处,南昌 330045)
昆虫生殖系统是内生细菌生长、发生和繁殖的理想场所,在长期特定环境的进化过程中,昆虫生殖系统与内生细菌之间相互选择、相互适应,导致不同来源、不同性别、不同发育阶段昆虫生殖系统内生细菌属种存在差异(Huangetal., 2010)。在昆虫生殖系统中,不同内生细菌属种,具有不同生理功能。例如,豆蚜Acyrthosiphonpisum生殖系统共生菌Buchneraaphidicola能促进豆蚜的生长和繁殖(Douglas, 1996; Russelletal., 2013);地中海实蝇Ceratitiscapitata生殖系统中克雷伯氏菌Klebsiellaoxytoca能够延长成虫寿命,提高成虫交配能力(Beharetal., 2008);伯克氏菌属Burkholderia能够促进昆虫生殖系统卵黄生成素的沉积,提高昆虫的产卵能力(Leeetal., 2017)。因此,研究雌雄昆虫生殖系统内生细菌属种及内生细菌在性别间的差异,对进一步探索昆虫生殖系统内生细菌的功能,及其对昆虫生殖行为和生殖能力的调控有积极意义。
橘小实蝇Bactroceradorsalis(Hendel)属双翅目Diptera实蝇科Tephritidae,是美国、日本和韩国等国家的重要检疫性害虫(张彬等, 2008; Wanetal., 2011)。橘小实蝇寄主多,能够为害柑橘、芒果和杨桃等46科250多种果蔬作物(胡凯平等, 2019; Zidaetal., 2020);成虫营两性生殖,具有极强的繁殖和扩张能力,每雌可产卵400~1 800粒,每年可扩张50~100 km(Duycketal., 2004; Lietal., 2019)。橘小实蝇雌雄成虫生殖系统中的内生细菌存在差异,能够调控橘小实蝇种群的生殖行为和生殖能力,进而影响到种群在田间的发生、危害与扩张(Yaoetal., 2016)。目前橘小实蝇肠道内生细菌已经受到科研人员的广泛关注(Wangetal., 2011; Wangetal., 2014);而生殖系统内生细菌的研究相对较少。仅王莉莉(2011)研究了室内饲养的橘小实蝇生殖系统中的细菌群落,并在雌性成虫生殖系统中发现植生拉乌尔菌Raoultellaplanticola、产气肠杆菌Enterobacteraerogenes、日沟维肠杆菌E.gergoviae和河生肠杆菌E.amnigenus等10种可培养菌群;而Shietal.(2012)发现,橘小实蝇雌性生殖系统的5种内生细菌,包括土生拉乌尔菌R.terrigena、产酸克雷伯氏菌K.oxytoca和肺炎克雷伯菌K.pneumoniae等的代谢产物,对橘小实蝇成虫本身具有一定的引诱作用。这些研究为揭示橘小实蝇生殖系统内生细菌的群落构成,分析内生细菌的生理功能提供了信息。
然而,由于橘小实蝇生殖系统内生细菌群落受众多外界因素影响,致使特定环境条件下橘小实蝇生殖系统内生细菌的组成存在差异。本研究以来源于江西赣南脐橙基地、以脐橙为主要食料的橘小实蝇为研究对象,采用LB培养基,对橘小实蝇雌雄成虫生殖系统中的内生细菌进行分离培养,分析菌株16S rRNA基因扩增序列,明确橘小实蝇雌雄成虫生殖系统内生细菌的多样性与差异,为进一步研究橘小实蝇生殖系统内生细菌在生殖调控中的作用奠定基础。
1 材料与方法
1.1 采集橘小实蝇样本
橘小实蝇样本采集时间为2018年9月中旬,采集地点为江西赣南脐橙基地(东经114°00′~114°40′,北纬25°42′~26°01′)。采集橘小实蝇样本时,收集携带橘小实蝇卵和幼虫的赣南脐橙果实,带回江西农业大学昆虫实验室,在温度28℃±1℃、相对湿度60%~90%和自然光照条件下,用赣南脐橙果实果肉组织饲养;老熟幼虫转移到用高温消毒后的沙土中,化蛹;成虫用酵母粉、清水和浓度为15%蜂蜜稀释液饲养10 d,收集成虫置于-20℃冰箱中保存、备用。
1.2 提取生殖系统
选取无病变、品相良好的橘小实蝇雌雄成虫各10头,无菌水冲洗干净。在超净工作台内用75%酒精冲洗2次,每次2 min,倒掉酒精;用无菌水漂洗3次,去除杂质;再在加有少量无菌水的培养皿中解剖,取出雌雄成虫生殖系统,放入2 mL EP管中,加入1 mL无菌水后充分研磨混匀,用于内生细菌培养。雌雄成虫各重复3次。
1.3 培养生殖系统内生细菌
上述配制的菌液,用无菌水按照10-2~10-5进行梯度稀释,移液枪吸取50 μL,置于LB培养基上,用无菌涂布棒涂布均匀,静置10 min,将培养基移至32℃恒温培养箱内。培养2 d后,用接种环挑取生长良好的不同形态或颜色性状的单菌落,采用稀释涂布划线法,在提前准备好的经过灭菌消毒的LB培养基上进行划线培养,得到单一纯菌落。将纯化后的菌株转至NA培养基斜面上,置于4℃冰箱保存备用。各稀释菌液均重复3次。
1.4 观察菌落的表观特征
接种环挑取NA斜面上的纯菌株,再次接入LB固体培养基中培养5 d,观察记录菌落形态、颜色、隆起度、边缘形状、表面状态、光泽、干湿情况、透明度、细菌形状、芽孢有无和革兰氏染色等特征。
1.5 扩增16S rRNA基因片段
无菌条件下,挑取NA斜面上的纯菌株,接入1 mL高压灭过菌的NB液态培养基中,在32℃、180 r/min恒温振荡培养箱中振荡培养3 d。菌液PCR扩增反应的上游细菌通用引物27F(5′-AGAGTTTGATCCTGGCTCAG-3′)和下游细菌通用引物1492R(5′-TACGACTTAACCCCAATCGC-3′),均由上海生物工程股份技术有限公司合成。PCR反应体系共30 μL:10×PCR bufferⅡ(with Mg2+)3.0 μL,dNTP Mixture 0.5 μL,968GC和L1401各0.5 μL,菌液1 μL,TaKaRaLaTaq酶(5 U/μL)0.5 μL,ddH2O 24.0 μL。PCR反应条件:95℃预变性5 min,94℃变性30 s,56℃退火30 s,72℃复性1 min;循环30次,72℃延伸10 min,4℃保温。扩增的PCR产物送美因基因公司检测并测序。
1.6 分析可培养细菌属种
利用DNAstar软件对得到的序列进行编辑、整理,然后在NCBI GenBank数据库中进行BLAST数据库检索和同源性比对,找出同源性最高的序列。利用Clustal X软件对测序获得的16S rRNA基因序列和与之近缘的已知序列进行多序性匹配排列,确定测试序列代表的细菌分类地位(Saitou and Nei, 1987; Thompsonetal., 1997)。
2 结果与分析
2.1 可培养细菌的菌落特征
采用LB培养基,从饲养10 d的橘小实蝇雌雄成虫生殖系统中,培养出具有不同形态或颜色性状的细菌15株。其中,雌性成虫9株,雄性成虫6株。不同菌株菌落,颜色多变,但多为白色、灰白色、黄白色或黄色。革兰氏染色表明,15株细菌菌落中11株呈革兰氏染色阴性反应,4株呈革兰氏染色阳性反应。显微镜观察显示,13株细菌呈杆状,2种呈球状(表1)。这些细菌均可以在32℃条件下生长,培养2~3 d后菌落形态、颜色性状明显。
2.2 可培养细菌属种鉴定
以橘小实蝇雌雄成虫生殖系统分离培养出来的15株细菌,分别配制的菌液为模板,对菌株16S rRNA基因进行扩增。扩增产物送美因基因公司检测、测序,比对鉴定可培养菌株属种。
测得的序列提交到NCBI进行blast同源性比对,发现橘小实蝇雌性生殖系统可培养的9株细菌来自4门、9属。变形菌门Proteobacteria内有4属,不动杆菌属Acinetobacter,苍白杆菌属Ochrobactrum、无色杆菌属Achromobacter和剑菌属Ensifer;拟杆菌门Bacteroidetes内有2属,金黄杆菌属Chryseobacterium和鞘脂杆菌属Sphingobacterium;放线菌门Actinobacteria内有2属,微杆菌属Microbacterium和细球菌属Micrococcus;厚壁菌门Firmicutes内1属,芽孢杆菌属Bacillus。橘小实蝇雄性成虫生殖系统可培养的6株细菌来自3门、6属。变形菌门内有3属,不动杆菌属、苍白杆菌属和假单胞菌属;拟杆菌门内2属,金黄杆菌属和拟杆菌属Bacteroides;厚壁菌门1属,芽孢杆菌属(表2)。
对比分析表明,橘小实蝇雌雄成虫生殖系统共有4个相同的细菌属,分别是不动杆菌属、苍白杆菌属、金黄杆菌属和芽孢杆菌属。雌性成虫生殖系统特有的细菌属,无色杆菌属、剑菌属、鞘脂杆菌属、微杆菌属和细球菌属;雄性成虫生殖系统特有的细菌属,假单胞菌属和拟杆菌属(表2)。


表2 橘小实蝇雌雄成虫生殖系统的可培养细菌属种
2.3 可培养的优势细菌属
橘小实蝇雌雄成虫生殖系统可培养细菌各属菌株数量所占比例不同。变形菌门内的不动杆菌属菌株所占比例最高,分别占雌雄成虫生殖系统菌株数量的35.00%和31.58%,为可培养细菌的优势菌属。此外,在雌成虫生殖系统中,苍白杆菌属、无色杆菌属、拟杆菌属和鞘脂杆菌属所占比例较高;在雄成虫生殖系统中,微杆菌属和芽孢杆菌属所占比例较高(图1)。
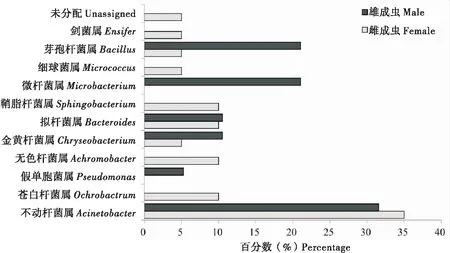
图1 橘小实蝇雌雄成虫生殖系统不同细菌属的比例
3 结论与讨论
本研究从橘小实蝇雌性成虫生殖系统中,分离培养出内生细菌9株,分别是不动杆菌属、苍白杆菌属、无色杆菌属、剑菌属、金黄杆菌属、鞘脂杆菌属、微杆菌属、细球菌属和芽孢杆菌属;从雄性成虫生殖系统中,分离培养出内生细菌6株,分别是不动杆菌属、苍白杆菌属、假单胞菌属、金黄杆菌属、拟杆菌属和芽孢杆菌属。这与王莉莉(2011)从橘小实蝇雌雄成虫生殖系统中分离培养出的内生细菌属种存在差异。可能原因,一是本研究种群来源于江西赣南脐橙基地,以脐橙为主要食料,饲养到成虫后直接研究细菌属种构成;而王莉莉(2011)的研究种群来源于广东出入境检验检疫部门,以香蕉为主要食料,且在室内饲养多代。二是分离培养条件不一致,虽然都采用了LB培养基,但培养基的制备、培养的温度和时间设置存在差异。三是测试方法亦存在差异,本研究通过扩增16S rRNA基因片段,然后分析鉴定细菌属种;而相关研究则采用高通量测序技术来分析橘小实蝇生殖系统细菌属种(王莉莉, 2011)。因此,研究材料来源、分离培养和测试方法的差异,决定本研究与前人的研究结果并不一样。
本研究结果表明,橘小实蝇雌性成虫生殖系统中的内生细菌门、属,均比雄性多。昆虫生殖系统中的内生细菌与昆虫的性别、发育阶段以及昆虫所处的外界环境条件密切相关(Beharetal., 2008; Sharonetal., 2010)。由于实验所用的橘小实蝇成虫的发育阶段一致且来自同一采样地点,具有相似的发育阶段和外界条件。因此,本研究橘小实蝇雌雄成虫生殖系统内生细菌的属种变化,可能与橘小实蝇的性别有关。
橘小实蝇雌雄成虫生殖系统均含有不动杆菌属、苍白杆菌属、金黄杆菌属和芽孢杆菌属,其中不动杆菌属为优势可培养内生细菌属。除橘小实蝇外,研究证实不动杆菌属在南瓜实蝇B.tau(骆米娟等, 2016)和按实蝇Anastrephaludens肠道(Lyudmilaetal., 2001)、泽兰实蝇Procecidocharesutilis幼虫体内(兰明先等, 2018)、番石榴实蝇B.orrecta成虫体内(戴阳等, 2020)等多种双翅目昆虫中具有分布。因此,可认为不动杆菌属在双翅目实蝇科昆虫中普遍存在。有报道,苍白杆菌属在番石榴实蝇成虫体内具有分布(戴阳等, 2020);芽孢杆菌属在泽兰实蝇幼虫体内具有分布(兰明先等, 2018);但未查到相关文献,金黄杆菌属在相关实蝇昆虫体内具有分布。
橘小实蝇雌成虫特有的细菌有无色杆菌属、鞘脂杆菌属、微杆菌属、细球菌属和剑菌属。其中,无色杆菌属和细球菌属在瓜实蝇B.cucurbitae体内有分布(姚明燕等, 2017);鞘脂杆菌属、微杆菌属和细球菌属在多种鳞翅目昆虫棉铃虫Helicoverpaarmigera、稻纵卷叶螟Cnaphalocrocismedinalis、菜青虫Pierisrapae和甜菜夜蛾Spodopteraexergual肠道中发现,鞘脂杆菌属能够诱导细胞发生程序性死亡(杨焊, 2012);剑菌属多在植物尤其是豆科植物中广泛分布,与植物固氮作用有关(陈文峰, 2016),昆虫中未查到相关文献。雄成虫特有的细菌属有假单胞菌属和拟杆菌属。研究证实,假单胞菌属在墨西哥实蝇(Lyudmilaetal., 2001)和南瓜实蝇雄成虫(骆米娟等, 2016)体内亦有分布,能抵御有害病原真菌的侵入与危害(Indiragandhietal., 2007);拟杆菌属在100 Gy137Cs辐照后的瓜实蝇肠道亦有发现(姚明燕等, 2017)。至于这些特异性细菌属种在橘小实蝇雌雄成虫生殖系统中所起的作用,目前仍不清楚。
本研究从橘小实蝇雌雄成虫生殖系统,分离出不同性状的细菌15株,分属4门,11属,明确雌雄间内生细菌属种存在差异。仍需要从二方面进行研究,一方面是需要根据细菌属种的营养需求和生化特点,来设计针对性的培养基类型,从而分离纯化更多可培养的细菌属种;另一方面需要对分离纯化的各细菌属种的生化特点和生物学功能进行研究、探索。本研究对橘小实蝇雌雄成虫生殖系统可培养内生细菌属种的比较研究,为优化培养基质和培养条件、探索生殖系统相关细菌的代谢和功能提供信息。
猜你喜欢
今日畜牧兽医(2022年10期)2022-12-23
湖北植保(2022年5期)2022-10-20
四川蚕业(2020年2期)2020-12-18
福建林业科技(2020年2期)2020-06-23
医学信息(2018年20期)2018-12-28
中西医结合心血管病电子杂志(2018年26期)2018-12-12
新农村(2018年32期)2018-04-02
江苏农业科学(2017年13期)2017-09-28
Coco薇(2015年12期)2015-12-10
大学(2008年4期)2008-07-10
