
不同营养条件下甜菜夜蛾幼虫感染核型多角体病毒的敏感性及取食趋性
2021-10-20 06:42乔风娇邢龙生李长友郑桂玲
环境昆虫学报 2021年5期
乔风娇,邢龙生,李长友,郑桂玲,张 彬*
(1.青岛农业大学植物医学学院,山东省植物病虫害综合防控重点实验室,中澳农业与环境健康联合研究院,山东青岛 266109;2.岭南现代农业科学与技术广东省实验室深圳分中心,农业农村部农业基因数据分析重点实验室,中国农业科学院(深圳)农业基因组研究所,深圳 518120)
甜菜夜蛾Spodopteraexigua属鳞翅目Lepidoptera夜蛾科Noctuidae,可危害多种农业经济作物,是一种重要的世界性农业害虫(Behmer, 2009)。二十世纪八十年代以前,该虫在中国仅是一种偶发性害虫,很少造成危害。自1986年以来,甜菜夜蛾在我国爆发成灾的报道频发,目前已遍布全国大部分地区(文礼章等,2014)。多年来,一些地区不合理的农药施用致使甜菜夜蛾的抗性水平逐年提升,不但增加了防治成本,而且对环境造成了污染和破坏。国家倡导的农业“双减”政策和“绿色防控”措施对我国生态文明建设和食品安全具有重要的战略意义,其中以甜菜夜蛾核型多角体病毒(Spodopteraexiguamultiplenucleopolyhedrovirus, SeMNPV)为代表的生物防治在甜菜夜蛾的防治中发挥了重要作用,同时降低了化学农药对环境的污染(Chenetal., 2019)。
SeMNPV对甜菜夜蛾的防治效果会受到多种外部因素的影响。除了温湿度、光照等环境因素之外,昆虫自身的营养状态也是重要的影响因素。研究表明,营养是影响昆虫自身免疫力的重要因素之一(Simpson and Raubenheimer, 2001; Leeetal., 2005)。特别是蛋白质和碳水化合物的摄入对很多昆虫的生理表现具有至关重要的作用,并且对昆虫的新陈代谢及种群增长产生一定的影响(Efeyanetal., 2015)。另有研究表明,蛋白质是影响昆虫对病原微生物敏感性的关键成分(Alauxetal., 2010)。在非选择性实验中,感染后存活下来的虫体比对照组和感染后死亡的虫体内蛋白质含量更高,表明幼虫感染致死剂量的细菌后将会增加蛋白质的摄入量(Leeetal., 2005; Poveyetal., 2009)。此外,通过自主选择营养物质可以显著提高宿主昆虫对于病原微生物的抵抗力(Raubenheimer and Simpson, 2009)。糖类作为昆虫的主要能源物质,不仅为昆虫生命活动提供了主要能量来源,也为昆虫几丁质的合成和脂类的转化提供原料和前体物质(王延年,1984)。研究表明,只给冈比亚按蚊Anophelesgambiae喂食含糖量较低的血液时会显著降低其黑化免疫反应能力(Koellaetal., 2002; Schwartzetal., 2002)。
综上所述,不同营养结构对于昆虫抵御病原物侵染的能力具有重要影响,而且病原物侵染后的宿主昆虫的取食偏好性也会发生较大变化。不同地区的甜菜夜蛾田间种群营养结构存在较大差异,不同营养结构的甜菜夜蛾对于核多角体病毒的敏感性存在明显差异。研究表明,取食不同寄主的甜菜夜蛾在感染病毒后血淋巴中病毒增殖和血淋巴黑化率具明显差异,说明了饲料影响宿主的黑化反应从而导致病毒增殖量的不同(王金彦,2018)。通过观察取食不同食物的甜菜夜蛾感染病毒后中肠组织的病理变化,表明不同食物对病毒感染中肠组织的影响存在明显差异(郭玲等,2018)。然而,食物营养影响病毒感染昆虫体内的免疫响应机理尚不清楚,仍待进一步研究取证。本研究通过观察不同营养品系甜菜夜蛾幼虫对SeMNPV的敏感性,以及染毒前后甜菜夜蛾幼虫对不同营养饲料的取食趋性,评价不同营养条件下甜菜夜蛾应对SeMNPV侵染的敏感性及其取食行为的影响,研究结果将有助于进一步研究农田营养结构对害虫种群发生发展的影响。
1 材料与方法
1.1 材料
1.1.1供试病毒
SeMNPV美国株(SeMNPV-US1)由美国康奈尔大学Wang Ping博士提供。
1.1.2供试昆虫
甜菜夜蛾试验虫源采自青岛市城阳区上马镇大葱田(地理位置120.17°E,41.67°N)。在甜菜夜蛾人工饲料配方的基础上(肖伟等,2005),分别建立了高蛋白低糖(简称高蛋白)、中蛋白中糖(简称中蛋白)和低蛋白高糖(简称低蛋白)3个营养品系,并连续饲养多代作为本研究的供试昆虫。人工饲养条件为温度28℃±1℃,相对湿度60%±5%,光周期为L14∶D10。
1.1.3供试药品
酵母粉(赛默飞世尔科技有限公司),葡萄糖(天津博迪化工有限公司),琼脂粉(北京索莱宝科技有限公司),山梨酸钾(天津市光复精细化工研究所),对羟基苯甲酸甲酯(天津市光复精细化工研究所),维生素C(天津市福晨化学试剂厂),维生素B(天津市福晨化学试剂厂),肌醇(天津市光夏精细化工厂),胆固醇(国药集团化学试剂有限公司)。
1.2 方法
1.2.1人工饲料配制
本实验所用人工饲料配制方法基于前期相关研究(肖伟等,2005),通过改变饲料中酵母粉(P)和葡萄糖(C)的比例,设定配制了3种不同营养含量的人工饲料,用于建立3种营养品系的甜菜夜蛾:高蛋白(P∶C=7∶1),中蛋白(P∶C=1∶1),低蛋白(P∶C=1∶7)(表1)。
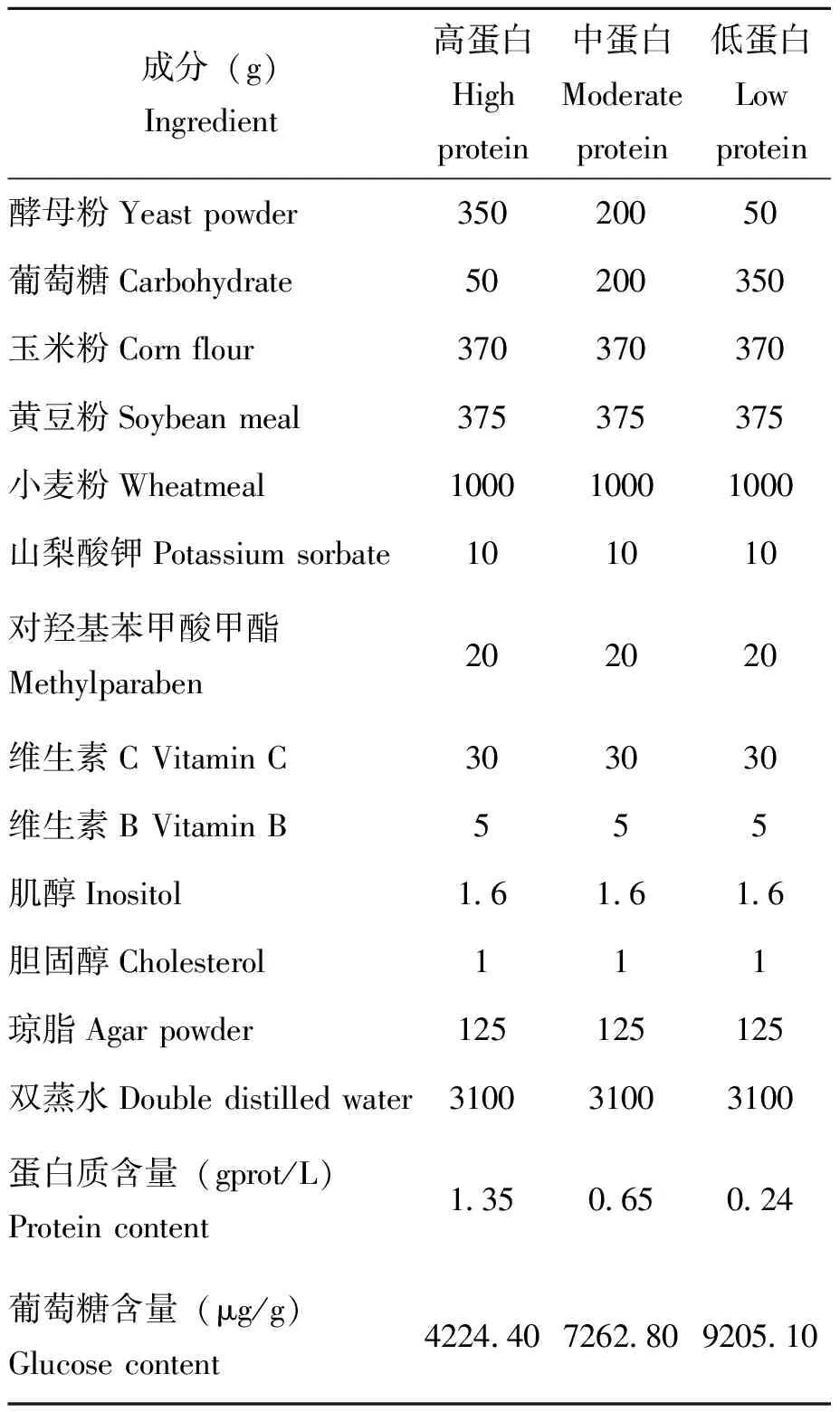
表1 不同营养饲料配制方法
1.2.2甜菜夜蛾初孵幼虫敏感性测定
参照文献中的生测方法(Hughesetal., 1986),在病毒悬液中加入藻蓝食用色素使最终浓度为0.5 mg/mL,测定甜菜夜蛾初孵幼虫对SeMNPV的敏感性。取SeMNPV-US1美国株多角体病毒,采用10倍梯度稀释,设8×102、8×103、8×104、8×105、8×106PIB/mL 5个处理组,以无菌水加入等量藻蓝食用色素作对照;每个浓度处理设置3重复,每个重复30头幼虫。
1.2.3染毒前后甜菜夜蛾幼虫取食趋性测定
用中蛋白中糖品系的甜菜夜蛾各龄期幼虫进行6 h饥饿后,分别设2组处理。一是在观察盒(直径6 cm高5.5 cm)内,两侧放置1 cm的高P低C、低P高C正方形饲料块,观察取食2种饲料虫数;二是将200 μL的8×106PIB/mL病毒悬液与16 g营养饲料混匀,得到病毒含量1×105PIB/g的饲料,用此饲料饲喂甜菜夜蛾幼虫6 h。然后将感染后的幼虫转入放置了高蛋白和低蛋白饲料的观察盒内,观察取食2种饲料的虫数。每个龄期处理设3个养虫盒,每盒10头幼虫,每天早晚各记录1次,未染毒与染毒幼虫趋性测定分别共进行3次重复。
1.2.4数据统计分析

2 结果与分析
2.1 不同营养品系甜菜夜蛾初孵幼虫对SeMNPV的敏感性差异分析
2.1.1SeMNPV对不同营养品系甜菜夜蛾初孵幼虫的LC50
为了明确病毒浓度对于初孵幼虫死亡率的影响,分别统计了不同病毒浓度条件下3种营养品系幼虫的累计校正死亡率。结果显示,随着病毒浓度的升高,3种品系幼虫的死亡率逐渐增大(图1)。当病毒浓度为8×105PIB/mL时,高蛋白、中蛋白质、低蛋白品系幼虫的累计校正死亡率依次为96.29%、79.02%、96.38%。此外,通过计算得到高蛋白、中蛋白、和低蛋白3种营养品系的LC50值依次为1.76×105、2.85×105、2.10×104PIB/mL(表2)。其中,中蛋白品系的LC50显著高于低蛋白品系(F=7.292,df=2,6,P=0.025<0.05),高蛋白与中蛋白、低蛋白品系均无显著性差异。上述结果表明,中蛋白品系对SeMNPV的敏感性最低。
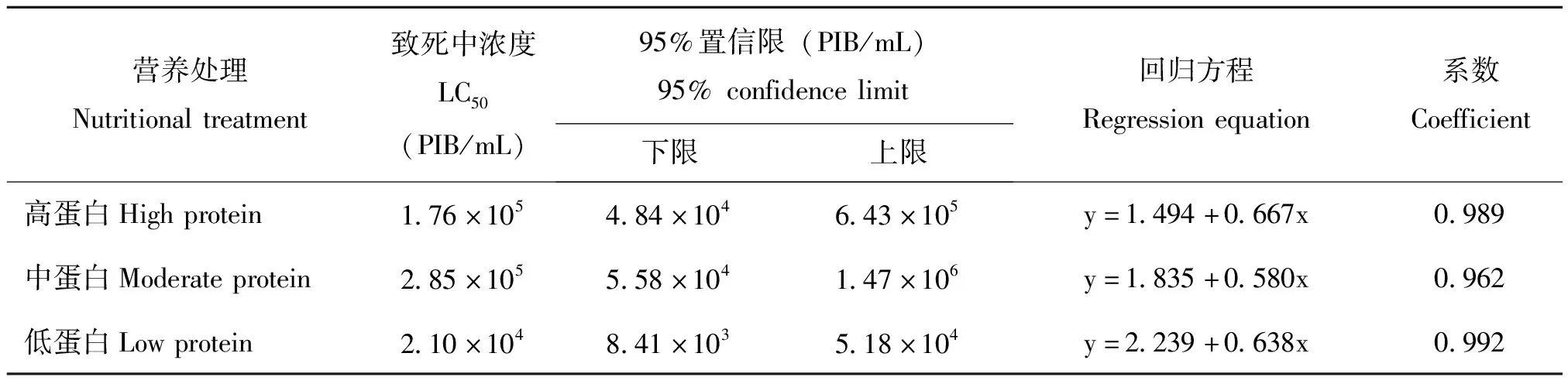
表2 不同营养条件下SeMNPV感染甜菜夜蛾初孵幼虫的LC50
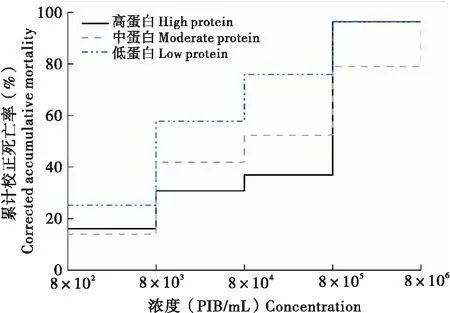
图1 不同营养条件下病毒浓度与甜菜夜蛾初孵幼虫的累计校正死亡率之间的关系
2.1.2SeMNPV对不同营养品系甜菜夜蛾初孵幼虫的LT50
3种营养品系幼虫第1~9天的累计校正死亡率随着感染病毒时间的增长逐渐增大(图2)。以浓度为8×105PIB/mL的病毒感染甜菜夜蛾幼虫时,高蛋白、中蛋白、低蛋白品系幼虫第9天的累计校正死亡率分别为96.29%、61.49%、96.42%。此外,通过计算得到3种营养品系的LT50值依次为6.07、6.85、5.20 d(表3)。统计分析表明,中蛋白品系的LT50显著高于低蛋白品系(F=8.927,df=2,6,P= 0.016 < 0.05),高蛋白品系的LT50同中蛋白品系以及低蛋白品系相比均无显著性差异。上述结果表明,中蛋白品系的甜菜夜蛾幼虫感染SeMNPV后存活时间最长。
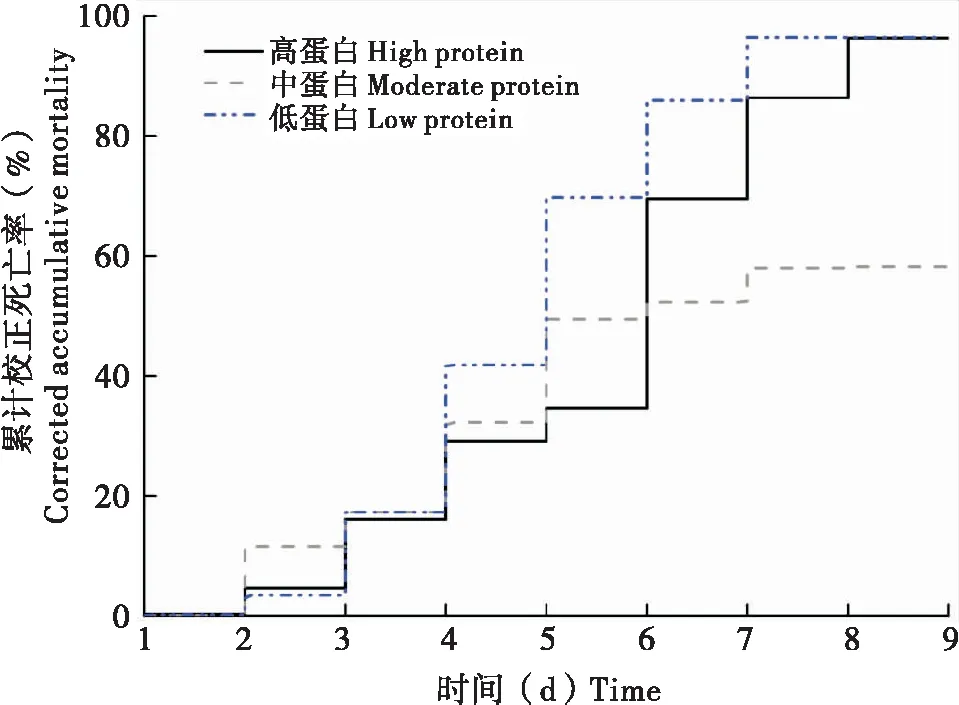
图2 3种营养品系甜菜夜蛾幼虫病毒侵染后的累计校正死亡率曲线
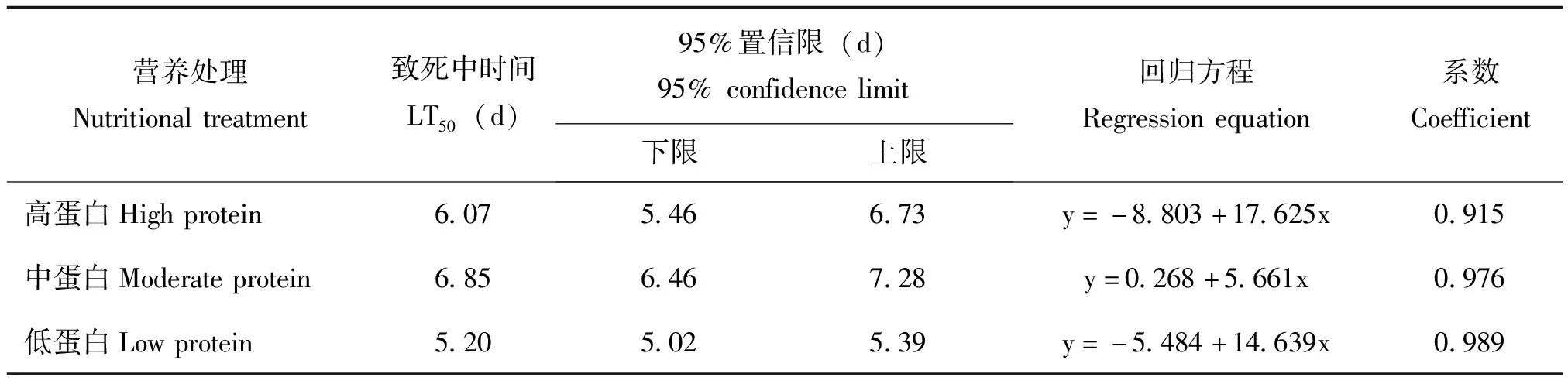
表3 不同营养条件下SeMNPV感染甜菜夜蛾初孵幼虫的LT50
2.2 甜菜夜蛾幼虫期取食趋性
2.2.1未感染SeMNPV幼虫的取食趋性
通过选择性实验发现,未感染病毒的幼虫在1~3龄幼虫取食高蛋白饲料显著多于取食低蛋白饲料的虫数(F=0.000,df=40.000,P<0.05),进入3龄末期之后,这种现象发生了显著变化,更多的幼虫偏向于取食低蛋白的饲料(F=0.000,df=28.000,P< 0.05)(图3)。
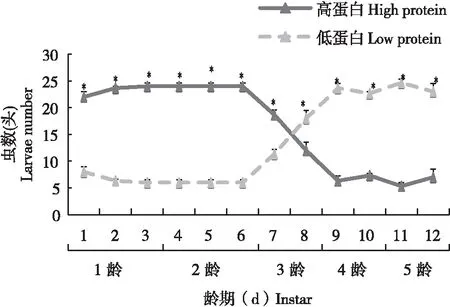
图3 未感染SeMNPV甜菜夜蛾幼虫对高蛋白和低蛋白饲料的选择
2.2.2感染SeMNPV幼虫的取食趋性
通过选择性实验发现,感染病毒之后的1~5龄幼虫均偏向于高蛋白的饲料。统计分析显示,各龄期幼虫取食高蛋白的饲料虫数显著多于取食低蛋白饲料的虫数(F=0.0000,df=70.000,P<0.05)(图4)。
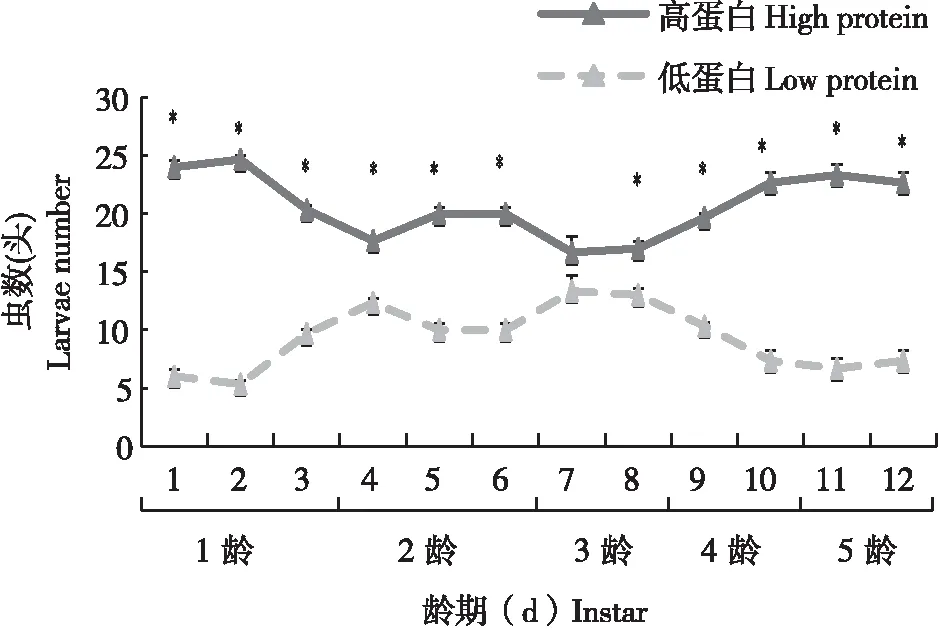
图4 感染SeMNPV甜菜夜蛾幼虫对高蛋白与低蛋白饲料的选择
3 结论与讨论
本研究通过比较不同营养条件下甜菜夜蛾初孵幼虫对核型多角体病毒侵染的敏感性差异,探究不同营养条件对昆虫抵御病原物侵染能力的影响。结果表明,中蛋白营养条件下幼虫对SeMNPV表现出较低的敏感性,说明适当的蛋白质和碳水化合物的比例对于甜菜夜蛾整个幼虫期对SeMNPV的抵抗力具有积极作用。相关研究表明,不同种类昆虫的最佳蛋白质和碳水化合物的比例存在差别,而且合适的蛋白质和碳水化合物的比例是提高昆虫免疫力的关键。本研究结果表明,饲料中合适蛋白质与碳水化合物的比例有助于甜菜夜蛾抵御病毒侵染,这一发现得到了前期相关研究结果的佐证。例如,东亚飞蝗Locustamigratoria喂食最佳配比的蛋白质与碳水化合物饮食后,在较高浓度的单宁酸下,昆虫的死亡率较低(Simpson and Raubenheimer, 2001)。另外,研究表明蛋白质与碳水化合物两种营养素会极大影响棉铃虫Helicoverpaarmigera(Hübner)对于毒素侵染的敏感性。同时,研究人员发现棉铃虫抵御病原物时自行选择P∶C饮食比为1.3∶1的食物(Tessnoweetal., 2017)。
通过对染毒前后的甜菜夜蛾幼虫对高P∶C比和低P∶C比两种人工饲料取食趋性的观察,本研究发现,未染毒情况下,甜菜夜蛾各龄期对营养需求表现出明显的龄期间的差异,基本表现为3龄之前偏向于取食高P∶C比的食料,3龄之后则偏向于取食低P∶C比的食料。这与前人报道一致(Chenetal., 2008),且符合以甜菜夜蛾为代表的鳞翅目农业害虫,低龄幼虫趋向取食蛋白质含量较高的植物幼嫩部位,而在高龄幼虫则可取食植物的成叶和老叶这些碳水化合物含量较高的部位的特征。但是,染毒后的甜菜夜蛾幼虫的取食偏好发生了显著变化。研究结果显示,染毒后甜菜夜蛾的低龄和高龄幼虫,都开始偏向于取食高P∶C比的饲料。前期研究报道指出,同低P∶C比饲料的甜菜夜蛾幼虫相比,高P∶C比饮食的甜菜夜蛾幼虫对SeMNPV的敏感性显著更低,表明摄取高蛋白的饲料更有利于增强宿主对SeMNPV侵染的抵抗力(Leeetal., 2005)。另有研究报道,蛋白质为昆虫免疫途径提供了必需的氨基酸,用于合成多种抗菌肽(Grimble, 2001; Schmid and Paul, 2005),表明当昆虫遭受病毒侵染时蛋白质是生物体调节体内免疫应答机制的重要营养物质。
综上所述,本研究结果明确了营养在甜菜夜蛾应对SeMNPV侵染过程中的重要作用,同时揭示了感染病毒前后各龄期幼虫取食偏好的差异性,为今后深入探讨营养调控甜菜夜蛾体内免疫应答的分子机制,提供了翔实可靠的分析结果和研究结论,也将有助于指导不同营养结构条件下的核多角体病毒的田间用药策略。
猜你喜欢
中国糖料(2022年2期)2022-04-06
云南农业(2021年11期)2021-11-12
小哥白尼(趣味科学)(2021年12期)2021-03-16
悦游 Condé Nast Traveler(2021年4期)2021-01-13
今日农业(2020年13期)2020-12-15
今日农业(2020年14期)2020-12-14
现代农村科技(2020年9期)2020-09-08
今日农业(2019年13期)2019-08-12
今日农业(2019年10期)2019-06-26
陕西农业科学(2019年3期)2019-04-09
