
海洋源生物活性肽的构效关系与作用机理研究进展
2021-10-31 11:25黄沐晨杨傅佳蔡茜茜吴金鸿汪少芸
食品科学 2021年19期
黄沐晨,杨傅佳,2,陈 旭,2,蔡茜茜,2,陈 选,2,吴金鸿,张 恒,张 军,汪少芸,
(1.福州大学生物科学与工程学院,福建 福州 350108;2.福州大学石油化工学院,福建 福州 350108;3.上海交通大学农业与生物学院,上海 200240;4.安徽国肽生物科技有限公司,安徽 宣城 242199;5.中食都庆(山东)生物技术有限公司,山东 菏泽 274108)
海洋是地球上最大的生物资源库。海洋生物链完整且再生能力强,其独特而复杂的孕育环境和悠久的演化发展历程,使海洋生物体内的防御体系和代谢机制具有特殊的性质[1],来自海洋生物的活性肽可分为内源性肽和外源性肽。内源性海洋生物活性肽是存在于海洋生物中的特异性功能肽,可积极发挥各种功能和生物学活性,主要包括海洋生命体中内分泌腺产生的肽类抗生素、激素等生物体代谢产物,血液或组织中存在的组织激肽以及神经递质或神经活动调节肽等[2]。可以运用分离纯化技术将内源性海洋生物活性肽直接从生命体中提取出来。外源性海洋生物活性肽是海洋源蛋白质通过水解产生的具有特定功效的肽,最主要的制备方法是酶解法[2]。根据不同酶的酶切位点可以产生多种不同功能活性的肽片段。通过微生物强大的酶系统发酵也是制备生物活性肽的有效手段,在发酵过程中,常选用枯草芽孢杆菌和真菌等从废弃的海洋源蛋白质中制备生物活性肽释放大量不同的二肽、三肽和其他生物活性肽[3]。化学合成法常用于已知氨基酸序列的海洋源生物活性肽,并可依据不同活性肽的构效机制对海洋源生物活性肽的氨基酸序列进行定向设计改造,从而快速获得理想有效的肽类药物[4]。根据生物活性肽的来源和种类,选择相应合适的制备方法以合理有效的获取目标活性肽尤为重要。
海洋生物是开发新型生物活性肽的良好资源,有着巨大的开发利用潜力[1]。随着现代生物技术和化学技术的发展应用,科技工作者们已利用丰富的海洋原料及其加工副产物,挖掘出众多结构新颖、功能特殊的生物活性肽,如抗氧化肽[5-8]、抗菌肽[9]、抗冻肽[10]、金属离子螯合肽[11-12]、免疫肽[13]等。近年来,有关海洋源生物活性肽的研究屡见报道。因此,有必要对海洋源生物活性肽的构效关系及作用机制进行综述,以期为海洋源生物活性肽的进一步挖掘及开发应用提供启发。
1 抗氧化肽
1.1 氨基酸序列与抗氧化的构效关系
自由基是机体代谢的中间产物,自由基的产生和积累会导致DNA的氧化损伤、蛋白质交联或降解、酶失活、发生脂质过氧化反应以及破坏细胞结构,造成机体免疫系统出现多种损伤和恶性病变,加速机体衰老,引发癌症、心血管疾病、阿尔茨海默症等慢性疾病的发展[5,8]。常见化学合成的抗氧化剂,如丁基化的羟基甲苯、叔丁基对苯二酚和丁基化的羟基茴香醚等存在毒性,危及人体健康。已有诸多研究表明,海洋加工副产物水解制备的抗氧化肽能够有效防止氧化造成的机体损伤,已成为最重要的人工抗氧化剂取代来源。
大多数抗氧化肽是由2~20 个氨基酸组成,低分子质量的肽相比于其亲本蛋白质更容易接近自由基从而抑制自由基介导的脂质过氧化。Kim等[14]发现抗氧化能力与肽的分子质量紧密相关,低分子质量组分(1~3 kDa)的鳕鱼蛋白水解产物组分相比于其他高分子质量组分具有更高的抗氧化活性。同样的,林琳等[15]发现以鱿鱼皮胶原蛋白为原料制备的小于2 kDa的多肽组分相比于其他分子质量更大的组分对羟自由基和超氧阴离子自由基的清除效果更好。Ranathunga等[16]发现,来源于康吉鳗的分子质量低于1 kDa的肽组分对体外脂质过氧化的防御效果优于天然抗氧化剂生育酚。
氨基酸组成及其位于肽序列中的位置对多肽抗氧化活性起着至关重要的作用。含苯环的氨基酸、酸性氨基酸和碱性氨基酸是抗氧化肽的活性位点,肽链的氨基酸位置决定了抗氧化肽到达细胞中自由基生成位点(如线粒体)的能力[17]。现有的研究发现抗氧化肽通常在N端含有疏水性氨基酸,增强了它们在脂质中的溶解度,使其易于进入靶器官发挥作用。颜阿娜等[18]从黑鲨鱼皮中酶解制备和分离纯化得到新型抗氧化肽Pro-Gly-Gly-Thr-Met(PGGTM),从图1中其清除1,1-二苯基-2-三硝基苯肼(1,1-diphenyl-2-picrylhydrazyl,DPPH)自由基机理图可以看出,N端的脯氨酸和C端的蛋氨酸可能是清除自由基的关键性位点。在抗氧化肽清除自由基的过程中,脯氨酸能够向DPPH自由基提供质子,并且两个带自由基的脯氨酸能够结合,从而阻止自由基链式传递[19-21]。疏水性氨基酸如蛋氨酸能增强多肽的抗氧化活性,蛋氨酸通过消耗周围环境的氧,被快速氧化生成甲硫氨酸亚砜,起到抗氧化作用[22]。Qian Zhongji等[23]采用体外胃肠消化法水解牡蛎得到抗氧化肽,确定了肽的氨基酸序列为LKQELEDLLEKQE(1.60 kDa),得到比生育酚活性更高的抗氧化肽。Yang Qian等[7]以黑鲨鱼皮为原料获得的新型抗氧化肽在500 μg/mL的质量浓度下对2,2’-联氮-双-3-乙基苯并噻唑啉-6-磺酸(2,2’-azinobis(3-ethylbenzothiazoline-6-sulfonic acid),ABTS)阳离子自由基具有81.5%的清除活性,序列为ATVY,其作用于ABTS的抗氧化作用位点为N末端的酪氨酸。Sinthusamran等[24]采用多种水解工艺酶解太平洋白对虾头胸部,不同的水解过程直接影响水解产物的蛋白质回收率、化学组成和抗氧化活性,得到的所有水解产物均以谷氨酸/谷氨酰胺、天冬氨酸/天冬酰胺、精氨酸和亮氨酸为主。另外,Zhao Kesheng等[25]发现Szeto Schiller(SS)肽可渗透细胞、靶向保护线粒体免受细胞氧化。SS肽的这种功能归因于它们有独特的结构基序,并含有芳香族和碱性氨基酸。

图1 Pro-Gly-Gly-Thr-Met清除DPPH自由基机理图[18]Fig. 1 Schematic illustration of the reaction mechanism between Pro-Gly-Gly-Thr-Met and 1,1-diphenyl-2-picrylhydrazyl radicals[18]
1.2 抗氧化机制
抗氧化肽在作用机理方面,主要从以下3 个方面抑制氧化反应[26]:1)通过抗氧化肽的肽键和羟基提供氢原子或电子清除自由基,以消除自由基对生物分子的破坏作用;2)通过钝化氧化酶的活性,螯合在氧化反应中起催化作用的金属离子,在脂质颗粒外形成保护膜等方式,封闭引起氧化的物质,阻截氧化反应的进行;3)上调内源性抗氧化剂如谷胱甘肽、超氧化物歧化酶和过氧化氢酶[27],调节细胞生理生化反应的抗氧化酶联防御功能[28],能够对细胞器破坏诱导慢性疾病发展起到抑制作用[29]。
抗氧化肽分子质量较小,容易穿过细胞膜被人体吸收利用,且安全性高,在吸收速率和生物学功能上具有显著优势。抗氧化肽可以用作食品配方中的成分,预防、控制和管理氧化应激相关疾病,还可以作为食品中的添加剂抑制食品的氧化。彭新颜等[30]发现蓝点马鲛鱼皮抗氧化肽可有效抑制4 ℃冷藏过程中熟肉糜蛋白降解和脂质氧化,保持熟猪肉糜品质。在保健应用及美白医药方面,王奕等[31]研究证实了日本刺参胶原肽对羟自由基等有显著的清除能力,并提高B16黑素瘤细胞的增殖水平,抑制酪氨酸酶活性进而抑制B16细胞黑素的合成。随着生命科学的发展和生物学作用机制研究的不断深入,抗氧化肽的构效关系将更加明确,海洋源抗氧化肽用于开发药物或作为保健品的功能因子将成为食品和高科技产业的前沿和热点,在医药领域将具有更好的发展前景和优势。
2 抗菌肽
2.1 二级结构与抗菌的构效关系
抗菌肽又称为肽类抗生素或抗微生物肽,是生物体内免疫防御机制经诱导产生的抵御外界微生物侵害的特殊小分子多肽。海洋源抗菌肽富含赖氨酸或精氨酸,使抗菌肽带正电荷,含较多的疏水性氨基酸形成疏水或两亲性结构[32],其分子质量小,容易到达被感染的位置,与传统抗生素不同,抗菌肽有广谱抗菌效果,且不会产生耐药性[33-34]。根据二级结构的差异可将抗菌肽分为4 类:α螺旋型、β折叠型、环形结构类以及伸展性螺旋结构类。
α螺旋型抗菌肽是水脂两亲性结构,疏水性氨基酸和带正电荷的亲水性氨基酸分布在螺旋轴的两侧,螺旋度的变化会影响其抗菌效果和穿透细菌细胞膜的能力[35]。β折叠型抗菌肽在氨基酸组成上富含半胱氨酸,可以形成分子内二硫键以稳定β折叠结构[35]。Qin Chuanli等[36]利用反相高效液相色谱从紫贻贝中分离出含55 个氨基酸残基的新型抗菌肽,这种新型抗菌肽对真菌和革兰氏阳性菌表现出卓越的抗菌活性,其分子质量为6 621.55 Da,具有一个几丁质结构域,分子中6 个半胱氨酸残基形成3 个分子内二硫键。环形结构类型在抗菌肽家族中较为少见,其环状结构由序列中的一个单键维持,这类抗菌肽可能同时具有α螺旋结构、β折叠结构。环形结构类型抗菌肽能对抗细菌和真菌,通过核磁共振(nuclear magnetic resonance,NMR)技术测定发现具有反平行β片层的结构,序列11位和18位间形成二硫键,通过二硫键稳定整个结构[35]。第4种抗菌肽为伸展性螺旋结构,其缺少典型的二级结构,一般呈线型,且氨基酸序列中不含半胱氨酸。富含脯氨酸的伸展性螺旋结构类抗菌肽,其结构含有多个Pro-Arg-Pro特征序列,部分抗菌肽在特定位置的氨基酸残基上还存在与生物功能密切相关的O-糖基化位点。
海洋源与陆生及两栖动物来源的抗菌肽在整体结构方面不具有显著的差别,但是,海洋源抗菌肽也存在一些如富含组氨酸的Piscidins这类特有的家族结构[37],为海洋源抗菌肽的开发应用提供了新思路。
2.2 膜损伤和非膜损伤机制
抗菌肽作为多细胞生物体内的防御物质,由于其不同于传统抗生素的作用机制而备受关注。对现有研究进行总结,抗菌肽的广谱抗菌作用主要分为膜损伤机制和非膜损伤机制。
膜损伤机制认为抗菌肽首先导致细胞壁结构变形,再通过静电相互作用导致细菌细胞质膜穿孔,破坏细胞膜产生孔隙,细胞内的水溶性物质外溢而死亡。破膜型抗菌肽的作用机制假说主要有图2所示的4 种[38]:桶板模型、环孔模型、毯式模型、聚集模型。在抗菌肽破膜模型中,当膜上抗菌肽的浓度超过阈值时便会插入细胞膜内,形成孔隙,即桶板模型。抗菌肽覆盖在膜周围,裂解膜形成地毯模型或插入到膜内形成环孔模型。聚集模型与去污剂作用模式相似,即结合于脂质膜表面的抗菌肽通过改变其自身结构与磷脂分子形成凝聚物,破坏膜结构,形成穿孔跨越细胞膜。Zhu Xin等[39]通过在α螺旋结构的氢键连接处放置不同类型的氨基酸残基来表征其抗微生物特性和作用机理,设计了一系列的两亲性肽,发现优化的序列通过诱导细胞质膜电位损失和改变膜通透性而表现出优异的抗微生物能力。海洋鱼类独有的抗菌肽Piscidins家族的作用机理属于典型的环孔模型机制。有研究者发现Piscidins-1相比于同源的Piscidins-3具有更强的杀菌能力,可能是由于N端2位的苯丙氨酸增强了螺旋结构的稳定性,而非极性部分17位的组氨酸则提高了破膜能力,改变了细胞膜的电导率和黏弹性,进而起到抗菌效果[40-41]。这一研究清楚地解释了Piscidins的结构与其抗菌作用机制间的关系。
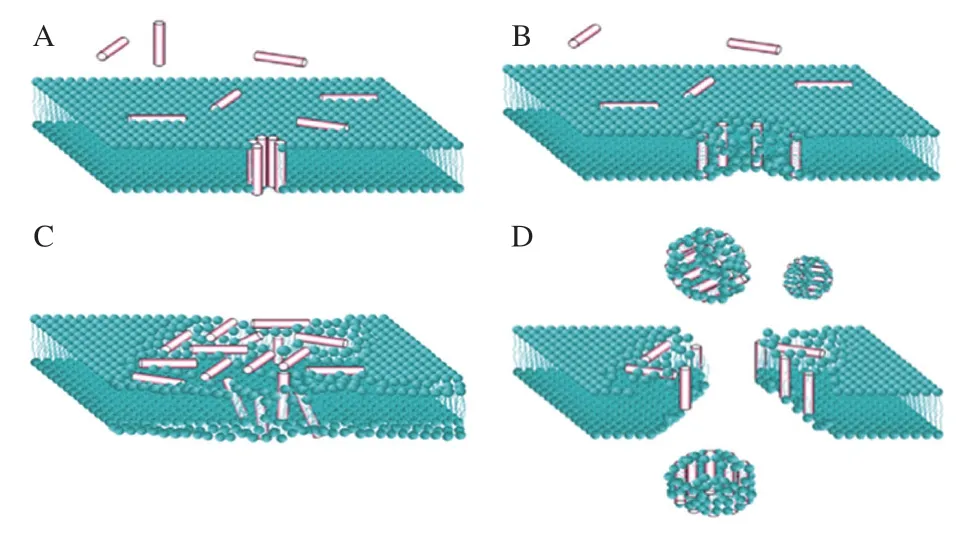
图2 文献中常被引用的抗菌肽作用模式图[38]Fig. 2 Modes of action of antimicrobial peptides against microbial membrane commonly reported in the literature[38]
非膜损伤机制通常作为一种辅助性作用方式,与细胞膜损伤机制联合作用于菌体细胞。部分抗菌肽可以穿过细胞壁膜进入细胞,作用于菌体胞内的靶点。如图3所示,抗菌肽的非膜损伤机制主要包括:抑制细胞壁合成,改变细胞脂膜(抑制细胞膜间隔的形成),诱导菌体自溶,抑制胞内酶活力,抑制DNA、RNA、蛋白质的合成以及与DNA结合等[42]。Powers等[43]研究者使用1H-NMR法测定来源于鲎的鲎肽I的结构,发现鲎肽 I是通过I型β转角连接的两亲性β-发夹结构,可转移到细胞中发挥抗菌作用。

图3 抗菌肽细胞内作用机制[42]Fig. 3 Intracellular mode of action of antimicrobial peptides[42]
抗菌肽除了在医药领域有望替代抗生素外,在食品行业也具有潜在的开发和利用价值[33]。宋宏霞[44]采用紫贻贝抗菌肽溶液浸泡草莓及鲈鱼,监测在一定贮藏条件下有关指标的变化情况,研究结果表明抗菌肽具有保鲜防腐的作用。海洋无脊椎动物只依靠体内免疫机制抵抗入侵细菌或病原体,因此内源性抗菌肽丰富[45],但海洋抗菌肽的获取存在提取操作繁琐、纯化成本高等问题,使其生产应用受限。提高抗菌肽的生产效率、降低成本是研究者们关注的焦点,高效经济的生产安全、稳定的海洋源抗菌肽对其发展和应用起有力的推动作用。
3 抗冻肽
3.1 冰晶-肽链的构效关系
抗冻蛋白(antifreeze protein,AFP)是一类具有热滞活性、能够抑制冰晶生长和重结晶的蛋白质,许多极地海洋鱼类能够存活于低温环境中正是得益于体内的AFP有效阻止血液在低温条件下凝固成冰[46]。海洋鱼源AFP根据氨基酸组成和结构不同可分为6 种[47-49](图4):抗冻糖蛋白(antifreeze glycoprotein,AFGPs)、AFP I、AFP II、AFP III和AFP IV及高活性AFP。AFGPs是最早被提取的AFP,肽链由肽糖单元重复连接而成,且发现糖基团是重要的抗冻活性基团[50],AFGPs在溶液中自发形成左手螺旋构像,保证肽链中的亲水的双糖基一侧朝向溶液,形成氢键与水分子结合。AFP I的分子质量为3.3~4.5 kDa,富含丙氨酸[51],在AFP中结构最为简单,其二级结构均呈现两亲性α螺旋,即亲水性氨基酸链与疏水性氨基酸链分别位于螺旋轴的两侧。AFP II结构特征表现为半胱氨酸含量较高,且约有一半能形成二硫键。研究证明,用巯基乙醇或者二硫苏糖醇破坏其二硫键会造成热滞活性丧失,因此,二硫键在AFP II分子中起着关键作用[52]。AFP III二级结构主要由9 个β折叠组成,其两两形成三明治样结构,剩余的游离在外,另外,AFP III中的亲水残基可以形成多个氢键使其和冰晶之间的结合力增强[53]。AFP IV的分子质量约为12.3 kDa,谷氨酰胺含量高,二级结构富含两亲性α螺旋,α螺旋以反向排列折叠成螺旋束的形式使得疏水基团朝内,亲水基团向外与冰晶结合[52]。Marshall等[54]从冬鲽鱼中分离得到一种高活性的AFP,它的活性和分子质量都比其他类型的AFP更高,使冬鲽鱼能够在-1.9 ℃的极限温度下生存。

图4 6 种鱼类AFP结构图[55]Fig. 4 Structural illustration of six kinds of fish antifreeze proteins[55]
但是,天然海洋源AFP含量甚微,应用成本高,随着对AFP活性结构研究的深入,科学家发现AFP的抗冻活性可来源于其一级结构多肽链中的某些片段[56]。海洋源抗冻肽则是利用海洋源蛋白质通过生物酶解技术,筛选出具有抗冻活性的肽段,一般具有GP-X(X为任何氨基酸残基)三肽重复序列、GTPG-和GPP(OH)_G-等特殊结构特征,且存在特定的氨基酸序列长度[10,57-59]。例如,Wang Shaoyun等[10]将鲨鱼皮胶原蛋白酶解产物进行分离纯化,得到了等电点小于5.0、分子质量为906 Da的纯化亲水性抗冻肽SsC-AFP,其氨基酸序列为GAIGPAGPLGP。对保加利亚乳杆菌进行低温胁迫处理,当SsC-AFP质量浓度为250 μg/mL时,菌种存活率高达90.28%。不同来源的抗冻肽无论结构或活性都存在着很大的差异,其抗冻机理、学说也不完全一致,阐明抗冻肽的作用机制对研究者来说仍然是一个巨大挑战。
3.2 吸附-抑制与亲水-互补作用机理
“吸附-抑制”学说认为AFP能够在冰晶结合面上吸附,形成冰曲面,通过开尔文效应抑制其生长[60]。在纯水体系中,冰晶垂直于晶体表面的方向生长(图5A),当AFP/抗冻肽分子吸附于冰生长路径的表面上(图5B、C),需要在外施加推动力促使冰在杂质间生长。具有特定分子质量和氨基酸结构的抗冻肽积累在冰-水界面,与水分子形成氢键吸附于冰核表面,改变冰晶生长方向和形态,形成六棱柱型或六角双锥体型,抑制冰晶生长及重结晶[10,57-59,61]。

图5 溶液中冰晶的形状[55]Fig. 5 Shapes of ice crystals in solutions[55]
汪少芸[10,57]、Damodaran[58]等根据活性肽氨基酸序列结构进一步对抗冻肽与冰结构层相互作用的理论模型进行了分子动力学模拟构建,采用表面“亲水-互补”模型对其作用机制进行分析[57],如图6所示,该模型认为,具有特定氨基酸长度和结构的抗冻胶原肽的冰晶结合位点所形成的表面与冰结构层面通过氢键互补联结,同时抗冻肽所含有的脯氨酸和丙氨酸残基等烷基侧链可以提供非极性环境以稳定氢键间的作用。抗冻肽与冰的结合阻止了冰晶的生长,表现出显著的冰晶生长抑制活性。
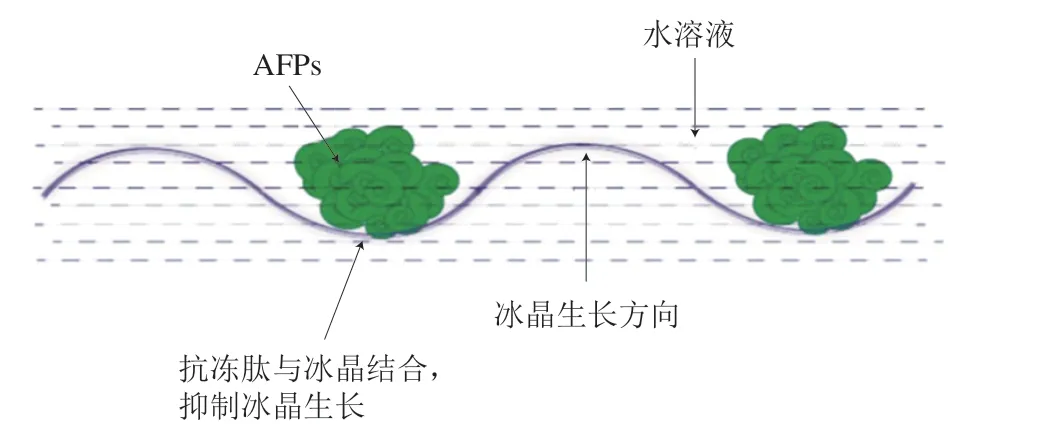
图6 抗冻多肽与冰晶表面亲水-互补作用示意图[62]Fig. 6 Surface hydrophilic-complementary interaction between antifreeze peptides and ice molecules[62]
抗冻肽具有良好抗冻活性的同时,也具有稳定的性质,获取手段便捷,制备量大,相较于AFP拥有更多的应用优势[56]。抗冻肽在食品工业中的应用主要体现在减少低温冷链食品的品质劣变,能够有效降低冷冻损伤,抑制冰晶形成、生长和重结晶。通过合理地开发利用海洋生物中丰富的蛋白质资源开发抗冻肽,替代现有高盐、高糖的食品抗冻剂或保水剂是一种新趋势。
4 金属离子螯合肽
4.1 肽-金属离子螯合的构效关系
金属元素对人体具有明显的生理功能和营养作用,它们的需求量小,却与机体生长代谢密切相关[63]。人体内对金属矿物元素吸收利用率较低,其缺乏会引起多种疾病,因此膳食金属元素补充剂的开发受到广泛关注。肽-金属离子螯合物可以利用机体对肽的吸收而使金属离子在消化道中更容易被吸收,具备优于无机态金属离子的生理生化特性。
影响金属螯合活性的结构特征包括肽的分子大小、氨基酸组成和特定的氨基酸序列,有研究指出海洋源金属离子螯合肽的羧基、氨基以及侧链中含孤对电子的氮、氧、硫原子,与金属阳离子通过化学键结合形成肽-金属离子螯合物[12,64]。Bao Xiaolan等[65]研究表明,某些特定氨基酸序列,如Asn-Cys-Ser被认为具有较高的螯合活性,特定氨基酸也能显著影响金属离子螯合肽的螯合作用,如天冬氨酸、谷氨酸和组氨酸,这些氨基酸上的羧基可能是螯合作用的结合位点,从而显著影响螯合反应。Zhang Lingtuo等[66]从裂殖壶菌粕蛋白水解产物中分离纯化出一种钙螯合肽,鉴定氨基酸序列为Ser-Ser-Val(SSV),分子质量为291.15 Da,具有良好的肽钙结合能力。利用傅里叶变换红外光谱分析发现该肽的螯合位点主要为羰基上的氧原子和氨基或亚氨基上的氮原子,SSV-Ca螯合物在酸碱条件下均表现出良好的热稳定性和溶解性,有利于人体胃肠道对钙的吸收和利用。Bouhallab等[67]研究发现,肽螯合金属离子的能力还与肽和金属离子结合的相对亲和常数、稳定常数等有关。此外,蛋白的磷酸化位点会影响肽-金属离子螯合反应,Wang Jiaxin等[68]发现酪蛋白的去磷酸化降低了酪蛋白磷酸肽与锌离子的螯合作用。多肽分子可以通过N端氨基、C端羧基、氨基酸侧链以及肽链中的羰基和亚氨基螯合金属离子,使其具有更高的配合率和稳定性,并且相较于其他类型的金属离子补充剂具有无毒副作用、制备成本低、操作便捷等优点[63]。
4.2 肽-金属离子螯合物的吸收作用机制
肽-金属离子螯合物结构稳定,肽对金属离子的保护可以避免金属离子受食物中的植酸和草酸的沉淀,从而提高其生物利用度[69]。
目前出现了两种被人们较为接受的肽-金属离子螯合物吸收机制的假说。1)完整吸收假说。即肽作为有机外壳将金属阳离子包裹起来,肽-金属离子螯合物以肽的形式被完整吸收进入小肠,通过肽的吸收通道,在小肠中进行转运[70],以完整的结构进入循环系统。2)竞争吸收假说。即多肽金属螯合物中的金属元素通过竞争方式被吸收利用,螯合物整体进入消化道后,金属元素受肽的包裹保护到达小肠黏膜的刷状缘,在吸收位点处肽-金属离子螯合物发生水解,膳食金属矿物元素分离出来,以离子形式进入肠上皮细胞,通过金属离子通道进入体内消化。Feng Yinuo等[71]发现酪蛋白水解物能够防止锌离子与磷酸钙发生聚沉,提高小鼠肠道对锌的吸收,推断Zn/CaP-纳米复合物一部分在细胞溶酶体中降解释放Zn2+到细胞质中;另一部分经内吞作用进入基底外侧(图7)。Ferraretto等[72]的研究进一步说明,螯合物的作用方式可能是肽直接嵌入到细胞膜中形成特殊的Ca2+选择性通道,作为一种钙载体活性肽经过细胞膜内吞作用使Ca2+进入细胞溶质中。凭借机体对肽类的吸收通路机制,肽与金属离子的螯合物具有作为一类新式金属离子增补剂的潜质,用于提升金属离子在机体的生物利用度和生物利用速度。
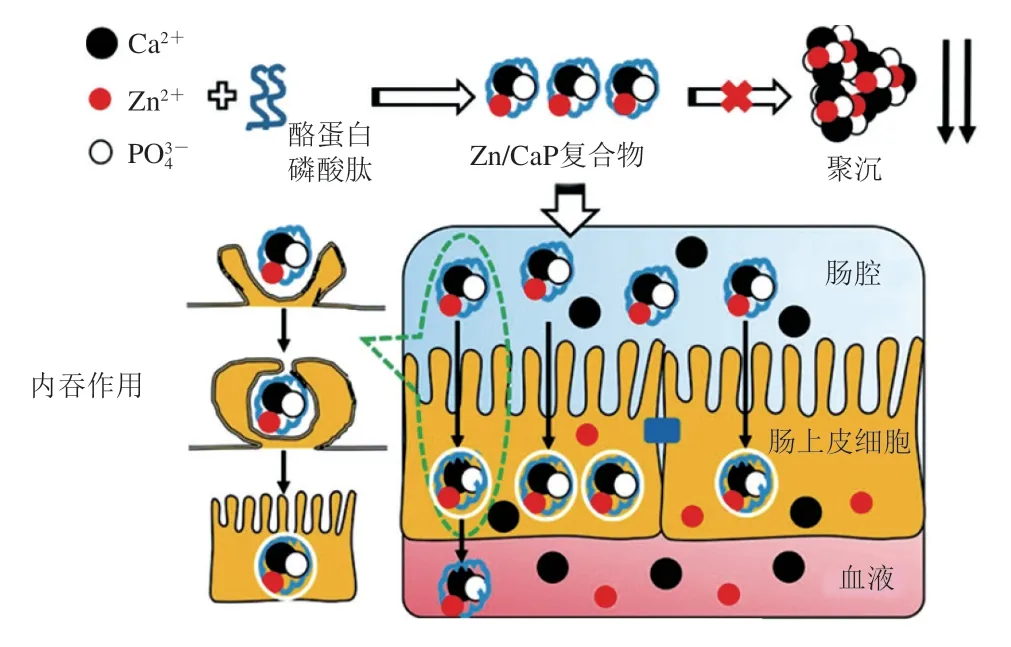
图7 Zn/CaP纳米复合物的肠吸收[71]Fig. 7 Intestinal absorption of Zn/CaP nanocomplexes[71]
目前,研究者们通过各种方式制备分离得到海洋源金属离子螯合肽,在与各类金属离子发生螯合作用后可以作为抑菌剂[73]、抗氧化剂[74]和人体金属离子补充剂[75]等产品应用到工业化生产中,具有无毒副作用、生物利用率更高、制备成本更为低廉、流程更为简便等优点,有着广泛的商业价值潜力和研究前景。
5 免疫调节肽
5.1 肽与免疫调节的构效关系
免疫力降低是人体代谢失常的根本原因,免疫系统是抵御病原体的第一道防线,能够在身体机能受损之前提供保护。因此,机体免疫功能的强化是疾病防控及诊疗的关键。海洋生物成为新型免疫调节肽提取的重要来源。Chalamaiah等[76]对不同酶水解产生的鱼卵蛋白水解物的体内免疫调节潜力进行了评估。在45 d内,对不同浓度的鱼卵蛋白水解物进行了处理。胃蛋白酶水解产物增加了脾巨噬细胞的细胞毒性和吞噬活性以及血清中免疫球蛋白A的水平。同时,胃蛋白酶和碱性蛋白酶水解物增强肠道内黏膜免疫,而胰蛋白酶水解产物增加了脾脏中CD4+、CD8+细胞数量。这些研究表明,水解产物的免疫调节作用与蛋白酶的类型有关,酶的特异性决定了生物活性肽的片段大小和序列分布。有研究报道,阿拉斯加鳕鱼的蛋白水解物能够刺激小鼠脾脏细胞的淋巴细胞增殖,与此活性相关的多肽的序列为NGMTY、NGLAP、WT[77]。免疫效应可能与氨基酸的物理化学特性相关,如正电荷数量、疏水性和链长。带正电荷的肽类似于趋化因子,可以结合免疫细胞上的受体,激活免疫反应。Yang Ruiyue等[78]在鲑鱼酶解产物中发现其氨基酸组成主要为Asp、Glu、Lys和Leu,得到的寡肽可以显著提高刀豆蛋白的活性、刺激淋巴细胞增殖、促进分泌细胞因子、调节自然杀伤细胞活性等。Merly等[79]在对鲨鱼软骨酶解产物的研究中发现,产物中Ⅱ型胶原蛋白α-1链和糖肽可进一步诱导机体的免疫反应。Wang Yukai等[80]对牡蛎水解多肽的免疫调节作用进行了研究,经牡蛎水解多肽处理的小鼠,体内淋巴细胞数量明显增多,同时发现自然杀伤细胞的活性也显著提高,血清中白细胞介素2分泌水平也有所提高。蛋白质被人体摄入后经过消化道酶解作用,多以小肽的形式通过小肠绒毛上皮细胞消化吸收。根据体内外的研究证据表明,免疫调节肽被认为是一种具有潜力的调节免疫反应策略物质,能够增强对感染因子的反应并减少组织损伤,对减轻自身免疫性疾病、炎症发生或与免疫系统相关的其他过程有着重要作用。
5.2 肽-特定细胞免疫调节的作用机制
免疫调节肽可通过调节淋巴细胞、单核细胞和粒细胞等刺激机体的免疫功能,关于免疫活性肽的调节机制一直是研究者们关注的热点[81]。外源性免疫调节肽可以通过直接作用于免疫系统的特定细胞发挥调节作用,其作用途径主要通过刺激特定细胞的细胞因子的分泌,诱导免疫细胞分化或激活免疫相关的细胞通路等[82-83]。
目前,研究者对免疫调节肽与免疫系统特定细胞之间的相互作用假说猜测与抗高血压和阿片肽系统有关[82-83]。如图8所示,相关研究表明免疫调节肽能够与位于免疫细胞表面的δ、μ型或κ型阿片受体相互作用,中枢阿片受体的激活可以调节周围的免疫系统[84-85]。酪啡肽在与阿片受体相互作用时,抑制了伴刀豆球蛋白刺激的淋巴细胞DNA合成和淋巴细胞中鸟氨酸脱羧酶的活性,从而抑制了体外固有层淋巴细胞增殖,影响人体黏膜免疫系统。肽序列决定了免疫细胞的反应活性,免疫调节肽的作用可归因于N端或C端区域的精氨酸、色氨酸或磷酸化的丝氨酸残基,以及谷氨酰胺的存在,这些氨基酸能够被免疫细胞表面的阿片受体识别[83]。

图8 肽与免疫细胞相互作用的相关机制[84]Fig. 8 Mechanism associated with peptides and their interactions with immune cells[84]
血管紧张素转化酶(angiotensin converting enzyme,ACE)抑制肽是高活性酪蛋白血管紧张素转化酶抑制肽,对于高血压有很好的治疗效果。免疫调节作用也与ACE抑制肽的作用机制相关。在ACE抑制肽发挥功效时形成的缓激肽是一种免疫调节肽,因此ACE抑制肽也具有免疫调节活性。缓激肽能够与靶细胞上的激肽受体结合,通过刺激巨噬细胞促进淋巴细胞增殖和转移,并诱导淋巴因子激活的杀伤细胞分泌细胞因子,发挥免疫调节作用[86]。免疫调节肽的N端或C端区域内有精氨酸可以被T细胞的膜结合受体识别,这些T细胞是淋巴因子的产生者。因此,C端精氨酸的存在是ACE抑制肽的结构特点,并且与免疫调节活性相关。
目前,许多药物在临床上用于控制人体免疫功能,包括左旋咪唑、咪喹莫特、匹多莫德等。但大多数合成的免疫调节药物都有一定的毒性和副作用,而天然来源的免疫调节蛋白或肽安全可靠且成本较低,现代营养学研究发现人类摄食蛋白质经消化道的酶解作用后,大多是以小肽的形式通过肠壁转运消化吸收,海洋源免疫调节肽可从多层面广泛地调节机体的免疫功能,对提高机体免疫力、确保机体健康具有重要的意义[87]。
6 结 语
此外,科研工作者发现海洋生物活性肽还具有抗衰老[88]、降血糖[89]、抗高血压[90]、ACE抑制活性[91]、抗炎[92]等功效,为此研究者还在不断地发现、分离、纯化新的海洋源功能肽类物质[1]。海洋源生物活性肽分子质量小,易被人体吸收和渗透到组织中;同时其毒副作用小、稳定性高,具有食疗效果等优点[93],这启发了研究者,海洋生物酶解提取物有望向功能食品的研发方向发展,同时充分利用海洋生物加工副产物,经过活性肽提取能够带来巨大的收益,提升商业价值。由于海洋资源的富饶,海洋环境的特殊和多样化,现有的活性功能肽作用机制有待深究,新型的海洋源生物活性肽也待科学家们去发现。
海洋源生物活性肽的开发仍然存在较大的局限性。酶解法是目前制备活性肽最为高效、普遍的方法,可以通过开发新型蛋白酶试剂、多种蛋白酶联合反应、采用多技术结合对酶解法获得海洋源生物活性肽进行深入优化,推进酶工程在海洋源生物活性肽领域的应用[9]。目前对海洋生物活性肽研究大部分仍处于实验室阶段,可以通过开发适用于产业化生产的活性肽分离设备,使得海洋生物活性肽能够从实验室科研水平进入产业规模化的生产。
海洋生物活性肽作为功能性食品及药品具有广阔的应用前景,但是在体内的研究还相对较少,在投入市场前,须对其活性进行检测和安全性评估[9]。此外,已有研究证明蛋白或肽的构象对这些分子的生物活性起到了决定性作用,因此,更加深入的构效关系分析极其必要,可以考虑对已经获得的海洋生物活性肽结构进行化学修饰,从而提高其活性和性能,有利于市场开发利用。
猜你喜欢
现代食品(2022年5期)2022-12-13
江西农业大学学报(2022年4期)2022-10-08
农林经济管理学报(2022年4期)2022-10-08
食品与生物技术学报(2021年5期)2021-06-24
医学概论(2021年19期)2021-01-21
潍坊学院学报(2020年6期)2020-11-22
世界科学技术-中医药现代化(2020年2期)2020-07-25
中成药(2019年12期)2020-01-04
中国食品学报(2019年10期)2019-11-12
中国油脂(2019年6期)2019-08-22
