
粉防己碱诱导雌激素受体阳性乳腺癌细胞自噬与凋亡的机制研究
2021-11-01 02:40赵立娜李以良祝东升
解放军医药杂志 2021年10期
赵立娜,翟 臻,梁 晨,李以良,李 巍,祝东升
雌激素受体(ER)阳性乳腺癌是最常见的乳腺癌亚型之一[1],其主要治疗方法为内分泌治疗,但长期治疗可能会导致多种不良反应及内分泌抵抗,从而降低了有效性[2]。因此,针对ER阳性乳腺癌的治疗方法仍旧面临重重困难,迫切需要开发新的治疗策略。
粉防己碱(tetrandrine,TET)是从粉防己(StephaniatetrandraS.Moore)根部提取的一种双苄基异喹啉生物碱[3]。多项研究已经证明TET通过诱导细胞凋亡从而发挥抗肿瘤活性[3-4]。内质网是细胞质内参与细胞稳态、凋亡和蛋白质折叠等多种生理过程的细胞器[5],内质网应激则是细胞自噬和(或)凋亡的重要途径[6]。基于上述理论研究,诱导内质网应激从而促进肿瘤细胞凋亡在抗癌治疗中的临床价值得到了越来越多的关注;但TET是否促进ER阳性乳腺癌细胞凋亡、是否通过内质网应激发挥促凋亡作用尚不清楚。本研究以BT474细胞为实验模型,揭示内质网应激对TET诱导ER阳性乳腺癌细胞自噬与凋亡的作用。
1 材料与方法
1.1细胞系 ER阳性乳腺癌细胞系BT474购自美国模式培养物研究所(ATCC)。
1.2主要试剂及仪器 TET(Sigma)、RPMI 1640(Gibco)、胎牛血清(FBA,Gibco)、胰酶(Gibco)、MTT(Sigma)、BCA蛋白定量试剂盒(Thermo)、Annexin V和PI凋亡试剂盒(索莱宝)。兔源抗LC3-Ⅱ单克隆抗体、兔源抗Beclin1单克隆抗体、兔源抗GRP78抗体、兔源抗p-PERK抗体、兔源抗p-eIF2α抗体、兔源抗CHOP抗体、兔源抗ATF6抗体、兔源抗β-actin抗体(Cell Signaling Technology,CST)、HRP标记羊抗兔IgG二抗(Santa Cruz Biotechnology)。酶标仪(Bio-Rad)、流式细胞仪(BD)、蛋白电泳系统(Bio-Rad)、凝胶成像系统(Bio-Rad)、细胞培养箱(Thermo)。
1.3细胞培养 乳腺癌细胞系BT474使用RPMI 1640 培养液+10% FBS,于37 ℃,5% CO2培养箱中培养。
1.4MTT检测 对数期BT474细胞经0.25%胰酶消化后,接种于96孔板中,每孔细胞数3×103个。培养24 h后,加入0(对照组)、5、15、30 μmol/L TET,于5% CO237 ℃培养箱中继续培养72 h。在培养终止前4 h,每孔沿孔壁加入5 mg/ml MTT溶液10 μl。培养结束后,小心吸弃培养液终止培养,每孔沿孔壁加入150 μl DMSO,轻柔震荡15 min。将96孔板放入酶标仪,检测490 nm处各孔吸光度。
1.5细胞凋亡检测 细胞以1×106个/孔的密度接种于6孔板中,用0、5、15、30 μmol/L TET处理72 h后收获细胞;PBS充分漂洗2次,沿孔壁加入200 μl缓冲液,轻弹6孔板底部以重悬细胞,分别吸取100 μl细胞悬浮液加入1.5 ml EP管中;根据细胞凋亡检测试剂盒(Thermo Fisher Scientific)说明书,取10 μl 1∶1混合的Annexin-V 和PI 预混液加入细胞悬浮液中,避光孵育15 min,加入150 μl结合缓冲液,采用FACScanⅡ流式细胞仪进行检测。计算凋亡细胞比例。
1.6蛋白免疫印迹法检测蛋白表达 收集0、5、15、30 μmol/L TET处理72 h后的BT474细胞,经RIPA裂解液裂解后,BCA蛋白定量检测蛋白含量,取40 μg蛋白上样检测Beclin1和LC3-Ⅱ蛋白含量。LC3-Ⅱ抗体(1∶2000)、Beclin1抗体(1∶2500)、GRP78抗体(1∶2000)、p-PERK抗体(1∶2500)、p-eIF2α抗体(1∶2500)、CHOP抗体(1∶5000)、ATF6抗体(1∶2000)、β-actin抗体(1∶2500),4 ℃孵育过夜;清洗后加入HRP标记羊抗兔IgG二抗,室温孵育1 h(1∶5000);清洗后参照BCL显色液说明书配置显色液,加入显色液至膜上;凝胶成像系统扫描成像,目标蛋白灰度值/内参灰度值代表相对蛋白表达量。

2 结果
2.1TET降低BT474细胞活力 TET 5、15、30 μmol/L组细胞活力分别下降10.5%、31.3%、57.2%。与对照组比较,TET 5、15、30 μmol/L组细胞活力显著降低,且呈浓度依赖性,差异有统计学意义(P<0.05,P<0.01)。见图1。
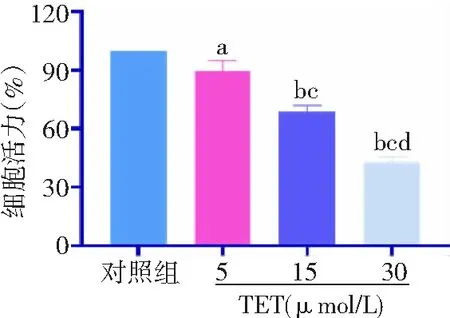
图1 不同浓度TET处理后BT474细胞活力比较对照组不加TET;TET为粉防己碱;与对照组比较,aP<0.05,bP<0.01;与5 μmol/L组比较,cP<0.01;与15 μmol/L组比较,dP<0.01
2.2TET促进BT474细胞凋亡 细胞凋亡率对照组为(10.35±0.78)%,TET 5、15和30 μmol/L组分别为(16.25±0.62)%、(33.49±0.82)%、(37.01±0.73)%。与对照组比较,TET 5、15和30 μmol/L组细胞凋亡率显著增高,且呈浓度依赖性,差异有统计学意义(P<0.05,P<0.01)。见图2。

图2 不同浓度TET处理后BT474细胞凋亡水平比较对照组不加TET;TET为粉防己碱;与对照组比较,aP<0.05,bP<0.01;与5 μmol/L组比较,cP<0.01;与15 μmol/L组比较,dP<0.01
2.3TET促进BT474细胞自噬 与对照组比较,TET 5、15、30 μmol/L组细胞自噬相关蛋白Beclin1和LC3-Ⅱ表达水平显著升高,且15、30 μmol/L组高于5 μmol/L 组,30 μmol/L组高于15 μmol/L组,差异有统计学意义(P<0.05,P<0.01)。图3、4。

图3 不同浓度TET处理后BT474细胞自噬相关蛋白Beclin1和LC3-Ⅱ表达蛋白免疫印迹图对照组不加TET;TET为粉防己碱

图4 不同浓度TET处理后BT474细胞自噬相关蛋白Beclin1和LC3-Ⅱ相对表达水平比较对照组不加TET;TET为粉防己碱;与对照组比较,aP<0.05,bP<0.01;与5 μmol/L组比较,cP<0.01;与15 μmol/L组比较,dP<0.01
2.4TET激活BT474细胞内质网应激 与对照组比较,TET 5、15、30 μmol/L组内质网应激通路相关蛋白萄萄糖调节蛋白78(GRP78)、p-PERK、p-eIF2α、CHOP表达水平升高,且15、30 μmol/L组高于5 μmol/L组,30 μmol/L组高于15 μmol/L组,差异有统计学意义(P<0.05,P<0.01)。对照组与TET 5 μmol/L组ATF6表达水平比较差异无统计学意义(P>0.05)。与对照组和5 μmol/L组比较,TET 15、30 μmol/L组ATF6表达水平升高,且30 μmol/L组高于15 μmol/L组(P<0.01)。见图5、6。
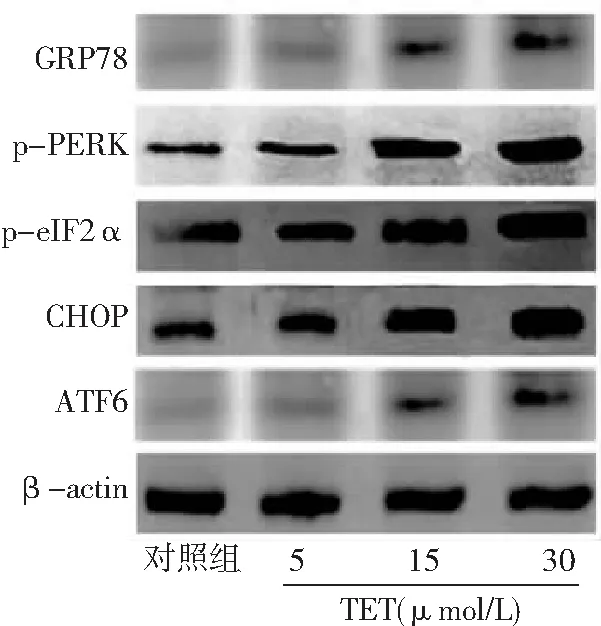
图5 不同浓度TET处理后BT474细胞内质网应激通路相关蛋白表达免疫印迹图对照组不加TET;TET为粉防己碱,GRP78为萄萄糖调节蛋白78
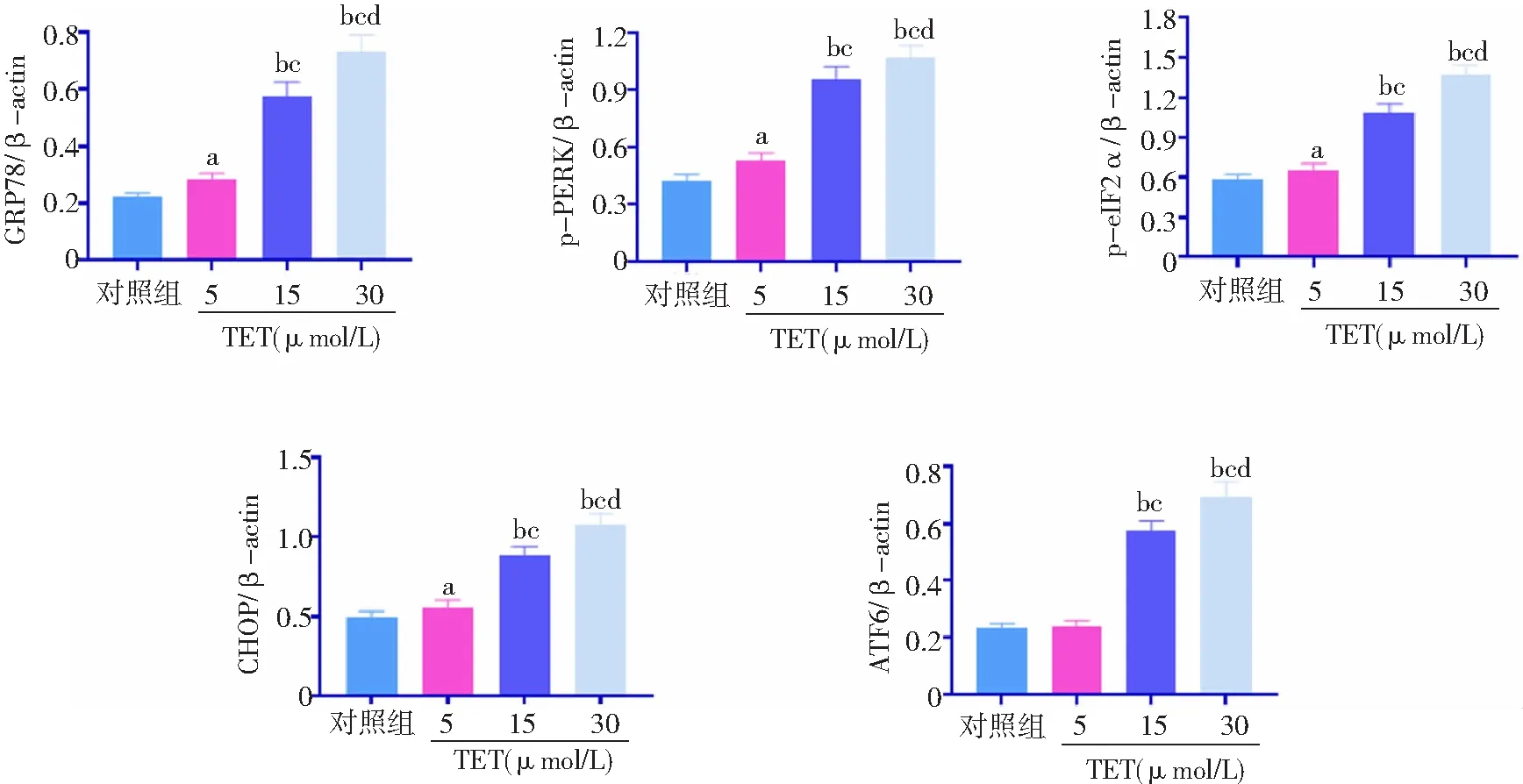
图6 不同浓度TET处理后BT474细胞内质网应激通路相关蛋白相对表达水平比较对照组不加TET;TET为粉防己碱,GRP78为萄萄糖调节蛋白78;与对照组比较,aP<0.05,bP<0.01;与5 μmol/L组比较,cP<0.01;与15 μmol/L组比较,dP<0.01
3 讨论
目前临床针对ER阳性乳腺癌治疗包括芳香化酶抑制剂和抗雌激素化合物,虽然内分泌治疗已经大幅降低了乳腺癌的复发率和病死率,但临床不良反应和获得性耐药仍然是该治疗方案的最大挑战[2]。因此,亟须探究和开发新的有效治疗手段。
TET对多种受体状态的乳腺癌细胞有抗增殖和促凋亡作用,增加放化疗的敏感性[6-7]。既往研究证实TET抑制PI3K/AKT/mTOR信号通路,从而促进三阴性乳腺癌细胞系MDA-MB-231的自噬和凋亡[3]。但TET是否促进ER阳性乳腺癌细胞自噬和凋亡及其相关作用机制研究较少。
自噬是溶酶体介导的细胞降解过程,该生理过程在肿瘤进展中起重要作用[8],也是癌症治疗的潜在靶点,是增强放疗和(或)化疗敏感性的策略之一[9]。Beclin1蛋白是自噬体形成的重要因子,介导自噬相关蛋白的亚细胞定位,从而调控自噬泡的形成及随后成熟[10]。LC3-Ⅱ附着于自噬体膜上,是自噬体的结构蛋白[11]。文献报道,TET诱导胃癌细胞自噬与凋亡,其作用机制与下调抗凋亡蛋白Bcl-2及促进细胞自噬标志物蛋白Beclin1表达有关[12]。本研究揭示了TET促进ER阳性乳腺癌细胞系BT474的自噬和凋亡,与既往研究一致,TET处理可提高BT474细胞中Beclin1和LC3-Ⅱ表达,证明TET能够激活自噬信号通路,从而抑制细胞增殖,促进细胞凋亡,且该效应具有明显的浓度依赖性。
内质网应激可通过激活细胞自噬信号通路导致细胞凋亡[13]。内质网应激是真核生物中重要的应激-防御机制之一,其通过未折叠蛋白反应(UPR)及其他相关分子信号调控自噬与细胞凋亡[13]。UPR信号通路由以下通路组成:PERK通路[14]、IRE1通路[15]和ATF6通路[16]。内质网分子伴侣GRP78是评估内质网应激水平的分子标志物[17]。Tiwari等[18]报道γ-生育三烯酚可提高乳腺癌细胞内质网应激和细胞自噬水平,促进癌细胞凋亡;同样,法卡地醇可通过内质网应激途径诱导癌细胞凋亡[19]。本研究发现TET可激活ER阳性乳腺癌细胞内质网应激信号通路相关蛋白GRP78、p-PERK、p-eIF2α、CHOP、ATF6表达,激活经典p-PERK、CHOP和ATF6通路。证实TET可能通过内质网应激激活细胞自噬细胞通路和细胞凋亡,从而抑制ER阳性乳腺癌细胞系BT474的增殖。与既往文献报道一致。本研究为TET治疗ER阳性乳腺癌提供了理论依据,但不足的是本研究使用的细胞系有限,后续可扩充乳腺癌细胞系类型以验证本结论。
综上所述,TET通过内质网应激通路激活细胞自噬,促进细胞凋亡,抑制ER阳性乳腺癌细胞BT474增殖。本研究丰富了TET在ER阳性乳腺癌中的作用机制,为临床的研究和应用奠定了基础。
猜你喜欢
现代临床医学(2021年1期)2021-01-26
亚热带植物科学(2020年5期)2020-12-25
亚太传统医药(2019年8期)2019-09-04
中成药(2018年12期)2018-12-29
安徽医科大学学报(2016年12期)2017-01-15
医学研究杂志(2015年4期)2015-06-10
——疾病防治的新靶标
中国药理学通报(2015年3期)2015-06-09
中国当代医药(2015年33期)2015-03-01
中国医药导报(2015年26期)2015-02-28
中华介入放射学电子杂志(2014年1期)2014-02-02
