
Exogenous platelet-derived growth factor improves neurovascular unit recovery after spinal cord injury
2021-11-02 02:59LuXiaYeNingChenAnPengHuangDuoHuiLiZhiLongZhengHaoJiHaoLiDaQingChenYanQingWuJianXiaoKeXuXiaoKunLiHongYuZhang
中国神经再生研究(英文版) 2021年4期
Lu-Xia Ye , Ning-Chen An , Peng Huang , Duo-Hui Li Zhi-Long Zheng Hao Ji,Hao Li, Da-Qing Chen, Yan-Qing Wu, Jian Xiao Ke Xu, , Xiao-Kun Li ,Hong-Yu Zhang
Abstract The blood-spinal cord barrier plays a vital role in recovery after spinal cord injury. The neurovascular unit concept emphasizes the relationship between nerves and vessels in the brain, while the effect of the blood-spinal cord barrier on the neurovascular unit is rarely reported in spinal cord injury studies. Mouse models of spinal cord injury were established by heavy object impact and then immediately injected with plateletderived growth factor (80 µg/kg) at the injury site. Our results showed that after platelet-derived growth factor administration, spinal cord injury, neuronal apoptosis, and blood-spinal cord barrier permeability were reduced, excessive astrocyte proliferation and the autophagyrelated apoptosis signaling pathway were inhibited, collagen synthesis was increased, and mouse locomotor function was improved. In vitro, human umbilical vein endothelial cells were established by exposure to 200 µM H2O2. At 2 hours prior to injury, in vitro cell models were treated with 5 ng/mL platelet-derived growth factor. Our results showed that expression of blood-spinal cord barrier-related proteins,including Occludin, Claudin 5, and β-catenin, was significantly decreased and autophagy was significantly reduced. Additionally, the protective effects of platelet-derived growth factor could be reversed by intraperitoneal injection of 80 mg/kg chloroquine, an autophagy inhibitor, for 3 successive days prior to spinal cord injury. Our findings suggest that platelet-derived growth factor can promote endothelial cell repair by regulating autophagy, improve the function of the blood-spinal cord barrier, and promote the recovery of locomotor function post-spinal cord injury. Approval for animal experiments was obtained from the Animal Ethics Committee, Wenzhou Medical University, China (approval No.wydw2018-0043) in July 2018.
Key Words: autophagy; blood-spinal cord barrier; central nervous system; locomotor function; neurovascular unit; platelet-derived growth factor; spinal cord; spinal cord injury
Introduction
Spinal cord injury (SCI) is a severe condition that may cause permanent disability (Ahuja et al., 2017; Courtine and Sofroniew, 2019), exhibiting primary and secondary injury patterns. The former comprises mechanical damage caused by local deformation of the spine, and the latter involves a complicated molecular reaction cascade (Mautes et al., 2000;McDonald and Sadowsky, 2002; Oyinbo, 2011; Telemacque et al., 2020). Among these pathological events, breakdown of the blood-spinal cord barrier (BSCB) is a critical mechanism leading to SCI progression and long-term neurological impairment (Whetstone et al., 2003; Lee et al., 2012; Koehn,2020). Numerous studies have demonstrated that functional recovery after SCI can be effectively promoted by preventing BSCB disruption (He et al., 2017; Zheng et al., 2017).Moreover, the relationship between autophagy and BSCB recovery in SCI remains unclear.
The neurovascular unit (NVU) is an interdependent multicellular complex composed of neurons, basal lamina,astrocytes, pericytes, and endothelial cells, in which metabolism of neurons and synapses is closely linked to capillary blood flow under astrocyte-mediated vasodilatory control (del Zoppo, 2010; Muoio et al., 2014; Iadecola, 2017).NVU pathophysiology is common in neurological diseases,with typical features including tissue hypoxia, inflammation,and angiogenic activation. Studies have indicated that intricate molecular interactions between cellular (brain endothelial cells, astrocytes, pericytes, and neurons) and acellular (basal lamina) components in the NVU can lead to enhanced bloodbrain barrier permeability and neurovascular uncoupling(Zlokovic, 2011; Stanimirovic and Friedman, 2012). Khan et al.(2009) reported that S-nitrosoglutathione administration in a rat model could protect the NVU and reduce secondary injury post-traumatic brain injury (TBI). Additionally, Ahmad et al.(2012) found that palmitoylethanolamide can protect the NVU and abate secondary injury after TBI in mice. Interestingly,the NVU concept is rarely reported in SCI. Epidermal growth factor, acidic fibroblast growth factor (FGF), and basic FGF have been shown to protect against blood-brain barrier or BSCB disruption post-TBI or -SCI (Wang et al., 2016; Zheng et al., 2016; Wu et al., 2017), indicating that the NVU may be of great significance in SCI and can be protected by growth factors.
As prototypes for growth factor and receptor tyrosine kinase function, platelet-derived growth factor (PDGF) and its receptor (PDGFR) have been studied for over 35 years(Keck et al., 1989; Battegay et al., 1994). PDGF and PDGFRs have clarified the role of PDGFR signaling in gastrulation and development of the skin, lungs, central nervous system,and skeleton (Hoch and Soriano, 2003; Andrae et al., 2008;Sun et al., 2020). Sufen et al. (2011) indicated that PDGF-BB can enhance the proliferation and migration of endothelial progenitor cells via the phospholipase C receptor signaling pathway. Additionally, PDGF-mediated autophagy can regulate vascular smooth muscle cell phenotypes and anti-oxidative stress responses (Salabei et al., 2013). In view of previous research on growth factors, including basic FGF (Zhang et al.,2013b), epidermal growth factor (Zheng et al., 2016), and FGF21 (Lu et al., 2019), and their participation in SCI and TBI through autophagy regulation, we hypothesized that PDGF can prevent disruption of the BSCB and improve NVU recovery by relieving SCI-induced autophagic flux damage. Therefore,we analyzed the effect of PDGF on SCI throughin vivoandin vitroexperiments.
Materials and Methods
Experimental animals and surgical procedures
Adult male C57BL/6 mice (specific pathogen free, 6–8 weeks old, 20–26 g) were obtained from Wenzhou Medical University(License No. SCXK [Zhe] 2015-0009). Mouse experiments were approved by the Ethics Committee of Wenzhou Medical University, China (approval No. wydw2018-0043) in July 2018 and were conducted in accordance with the guidelines of the National Institutes of Health. The animals were divided into sham, SCI, SCI + PDGF, SCI + chloroquine (CQ), and SCI + PDGF+ CQ groups (n= 6 per group). Mice were anesthetized with 4% isoflurane and placed on the platform. A skin incision was made along the midline of the back to expose the vertebral column, and a laminectomy was performed at the T9 level. A moderate contusion was generated by dropping a 10-g weight from a height of 2 cm (Liu et al., 2015). Then, the incision sites were closed in layers, and mice were allowed to recover from anesthesia. The bladders of mice were manually emptied twice daily.
Drug administration
PDGF-BB (100-14B; PeproTech, Rocky Hill, NJ, USA) and CQ (HY-17589; MCE, Monmouth Junction, NJ, USA) were administered in this study. PDGF stock solution was diluted with 0.9% NaCl and immediately injected once into the spinal cord after SCI (80 µg/kg). Animals were intraperitoneally injected with 80 mg/kg CQ daily for 3 days before injury.
Enzyme-linked immunosorbent assay assays
At 3 days after SCI, mice were anesthetized, and blood was collected from the eyelid. The serum was kept stationary and then assayed using a PDGF enzyme-linked immunosorbent assay protocol (Westang, Shanghai, China). Fluorescence was quantified with a spectrophotometer (Molecular Devices,Silicon Valley, CA, USA) at an excitation wavelength of 450 nm and an emission wavelength of 490 nm using a microplate reader (Molecular Devices).
Evans blue dye assays
BSCB penetrability was investigated via Evans blue dye. At 1 day after SCI, 2% Evans blue dye (2 mL/kg; Sigma-Aldrich,St. Louis, MO, USA) was injected into the tail vein of mice.Two hours later, animals were anesthetized with 4% (w/v)isoflurane and transcardially perfused with saline (0.9%NaCl). Then, 500 µL of N,N-dimethylformamide was added to the tissues, which were than incubated at 72°C for 3 days.Samples were centrifuged for 20 minutes. The supernatant(200 µL) was collected and added to 96-well glass plates.Fluorescence was quantified with a spectrophotometer at an excitation wavelength of 620 nm and an emission wavelength of 680 nm using a microplate reader. All experiments were repeated at least in triplicate.
Tissue preparation
Animals were anesthetized with 4% isoflurane and perfused by cardiac puncture with 0.9% saline at 3 and 14 days post-SCI. For immunofluorescence staining, 0.5 cm sections of dissected spinal cord lesions were obtained, followed by postfixation in 4% paraformaldehyde. Finally, paraffin embedded tissue sections (5 mm) were cut and placed onto slides for staining.
Western blot assay
After homogenization in radioimmunoprecipitation assay lysis buffer supplemented with a protease inhibitor cocktail (10µL/mL; GE Healthcare Biosciences, Pittsburgh, PA, USA), the homogenates were centrifuged to collect the supernatants. A bicinchoninic acid kit (Beyotime, Shanghai, China) was used to qualify the extracts. After separation and transfer, membranes were blocked in Tris-buffered saline-Tween-20 with 5% skim milk for 2 hours and incubated at 4°C overnight with primary antibodies (zonula occludens-1 [ZO-1; 1:1000; rabbit; Abcam,Cambridgeshire, UK], β-catenin [1:1000; rabbit; Abcam],Occludin [1:1000; rabbit; Abcam], Claudin 5 [1:1000; rabbit;Abcam], microtubule-associated proteins 1A/1B light chain 3B [LC3B; 1:1000; rabbit; Novus, Centennial, CO, USA], p62[1:1000; rabbit; Abcam], AKT [1:1000; rabbit; Abcam], p-AKT[1:1000; rabbit; Abcam], PDGFR [1:1000; rabbit; Abcam],phospho-(p-)PDGFR [1:1000; rabbit; Abcam], glyceraldehyde-3-phosphate dehydrogenase [1:1000; rabbit; Abcam], Bax[1:1000; rabbit; Cell Signaling Technology, Boston, MA,USA], and Bcl-2 [1:1000; rabbit; Cell Signaling Technology]),followed by incubation in horseradish peroxidase-conjugated goat anti-rabbit IgG (1:10,000; Cell Signaling Technology)and horseradish peroxidase-conjugated goat anti-mouse IgG (1:10,000; Cell Signaling Technology) for 1 hour at room temperature. Bands were imaged using a Chemi DocXRS+Imaging System (Bio-Rad, Berkeley, CA, USA), and the results were quantified by Quantity-One software (Bio-Rad).
Basso Mouse Scale
The recovery of hindlimb locomotor of mice at 1, 3, 7, 14, and 28 days post-SCI was assessed in an open field test using the Basso Mouse Scale (McTigue et al., 2007). The Basso Mouse Scale score ranged from 0 (complete paralysis) to 9 (normal movement of the hindlimbs) and was determined according to the average performance of the left and right hindlimbs.
Footprint analysis
A footprint analysis was performed at 14 days post-SCI by immersing the hind limb in a red dye, and the hind limb motion trajectory was printed on a white runway. The results were blindly recorded.
Pathological observation
Poly-L-lysine-coated slides containing paraffin sections (5 mm)were stained with hematoxylin and eosin. The experimental steps were carried out according to manufacturer’s instructions (Cat# C0105; Beyotime). For Nissl staining (Cat#C0117; Beyotime), sections were incubated in 1% cresyl violet and imaged with a light microscope. Masson staining was conducted according to the manufacturer’s instructions(Cat# G1340; Solarbio, Beijing, China). Regions of interest in the tissue sections were imaged using an optical microscope(Nikon, Tokyo, Japan).
Cell culture and treatment
Human umbilical vein endothelial cells were purchased from the Cell Bank of the Type Culture Collection of the Chinese Academy of Sciences, Shanghai Institute of Cell Biology,Chinese Academy of Sciences, cultivated in endothelial cell medium. and divided into control, H2O2, and H2O2+ PDGF groups. The cells were pre-treated with PDGF (5 ng/mL)for 2 hours before stimulation with 200 µM of H2O2. The H2O2concentration was based on our previous research report (Wang et al., 2018). Cell viability was assessed by Cell Counting Kit-8 solution (Beyotime) according to the manufacturer’s protocol. Pericytes (Shanghai Zhong Qiao Xin Zhou Biotechnology Co., Ltd., Shanghai, China) were added to 96-well plates, and the serum was depleted for 24 hours. The cells were then incubated with the indicated concentration of PDGF for 48 hours. Next, 10 µL of Cell Counting Kit-8 solution was added to each well, followed by incubation for 1 hour. The absorbance was measured at a wavelength of 450 nm using a microplate reader (Molecular Devices). Each experiment was performed in triplicate.
Immunofluorescence staining
Cells were fixed using 4% paraformaldehyde at room temperature. The cells and tissue sections were incubated with primary antibodies including anti-ZO-1 (1:1000; rabbit;Abcam), anti-LC3B (1:1000; rabbit; Abcam), anti-glial fibrillary acidic protein (GFAP; 1:1000; chicken; Abcam), anti-Lectin(1:1000; rabbit; Abcam), anti-β-Tubulin III (1:1000; rabbit;Abcam), and anti-NeuN (1:1000; mouse; Abcam) at 4°C overnight after blocking with 5% bovine serum albumin for 45 minutes, followed by incubation with Alexa Fluor fluorescein isothiocyanate, Alexa Fluor TRITC, or Alexa Fluor Cy5 goat anti-rabbit/mouse/chicken IgG (1:1000; Abcam) at room temperature for 1 hour. Next, the samples were treated with a fluorescence quenching solution including 4′,6-diamidino-2-phenylindole (Beyotime) and imaged under a microscope (A1 PLUS; Nikon). Ten images were randomly selected from the tissue samples, and ImageJ software (National Institutes of Health, Bethesda, MD, USA) was used for immunofluorescence quantification.
Statistical analysis
Data are expressed as the mean ± standard error of the mean (SEM). The statistical analysis was performed by oneway analysis of variance and Dunnett’spost hoctest using GraphPad Prism software version 8 (GraphPad Soft-ware Inc.,La Jolla, CA, USA). Statistical significance was set as aPvalue <0.05.
Results
PDGF reduces motor neuron loss and promotes functional recovery of SCI
To determine the role of PDGF in SCI recovery, the impact of acute SCI on PDGF protein expression in spinal cord tissue was assessed at 3 days after injury. PDGF was significantly augmented after acute SCI (P< 0.01; Figure 1A), similar to the findings observed by Chen et al. (2017), which revealed that PDGF might affect SCI. Therefore, PDGF was injected near the wound surface in mice to detect its efficacy in SCI treatment.Figure 1B shows obvious severe tissue loss in the SCI group compared with the sham group, while PDGF administration abated tissue damage and neuronal apoptosis. Furthermore,we hypothesized that PDGF had a protective effect on functional recovery after SCI. To verify this hypothesis, the Basso Mouse Scale scores were determined. The sham group scored an average of 9 points, indicating normal motor function, and the SCI + PDGF group exhibited improved functional recovery after 3 days (Figure 1C). In addition, we examined axonal regeneration after SCI. As shown in Figure 1D and E, axonal regeneration was improved in the PDGF group compared with the SCI group (P< 0.05). These data revealed that PDGF could reduce tissue damage, protect neurons, and ameliorate the pathological morphology of the lesion area after SCI.
The protective efficacy of PDGF on SCI-induced BSCB breakdown is regulated by activated tight junction and adherens junction proteins in vivo and in vitro
Tight junctions (TJs) and adherens junctions (AJs) are the structure and foundation of the BSCB, and changes in BSCB function are associated with the expression of these proteins(Ye et al., 2016; Lochhead et al., 2017). The Evans blue assay results suggested that BSCB permeability was enhanced in the SCI group compared with the control group, whereas Evans blue intake was decreased after PDGF treatment (P< 0.01;Figure 2A and B). Next, we evaluated the effect of PDGF on AJ protein (β-catenin) and TJ protein (Claudin 5, Occludin,and ZO-1) activity. An immunofluorescence assay showed that ZO-1 was markedly enhanced after PDGF administration compared with SCI mice (P< 0.01; Figure 2C and D). As shown in Figure 2E and F, PDGF upregulated TJ and AJ protein expression compared with the SCI group. To further verify that PDGF protected against BSCB breakdown, we examined the expression of BSCB-related proteinsin vitro. The expression levels of Occludin, Claudin 5, and β-catenin in the H2O2-treated cell group were significantly decreased, while PDGF increased the expression of these proteins (Figure 2G–J). These data suggested that PDGF prevented disruption of BSCB integrity after SCI.
PDGF protects the NVU after SCI in mice
Lectin, GFAP, and NeuN staining were performed to observe the morphologies of vessels, astrocytes, and neurons,respectively, in the injury area. As shown in Figure 3A and B,more vessels and neurons were present in the injury areas in the SCI + PDGF group than in the SCI group. Additionally, the number of astrocytes decreased with increases in the vascular network (Figure 3C and D), leading to the formation of glial scars. Moreover, PDGF could remodel the microenvironment in the NVU. As shown in Figure 3E, the PDGF group exhibited a significantly higher collagen level than the SCI group. Overall,these findings implied that PDGF could protect the whole NVU by reducing neuronal apoptosis, inhibiting of the excessive proliferation of astrocytes, and promoting collagen synthesis,which might be related to the promotion of vascularization(Figure 3F).

Figure 1 |Platelet-derived growth factor (PDGF) treatment decreases tissue damage and enhances functional recovery post-spinal cord injury (SCI).
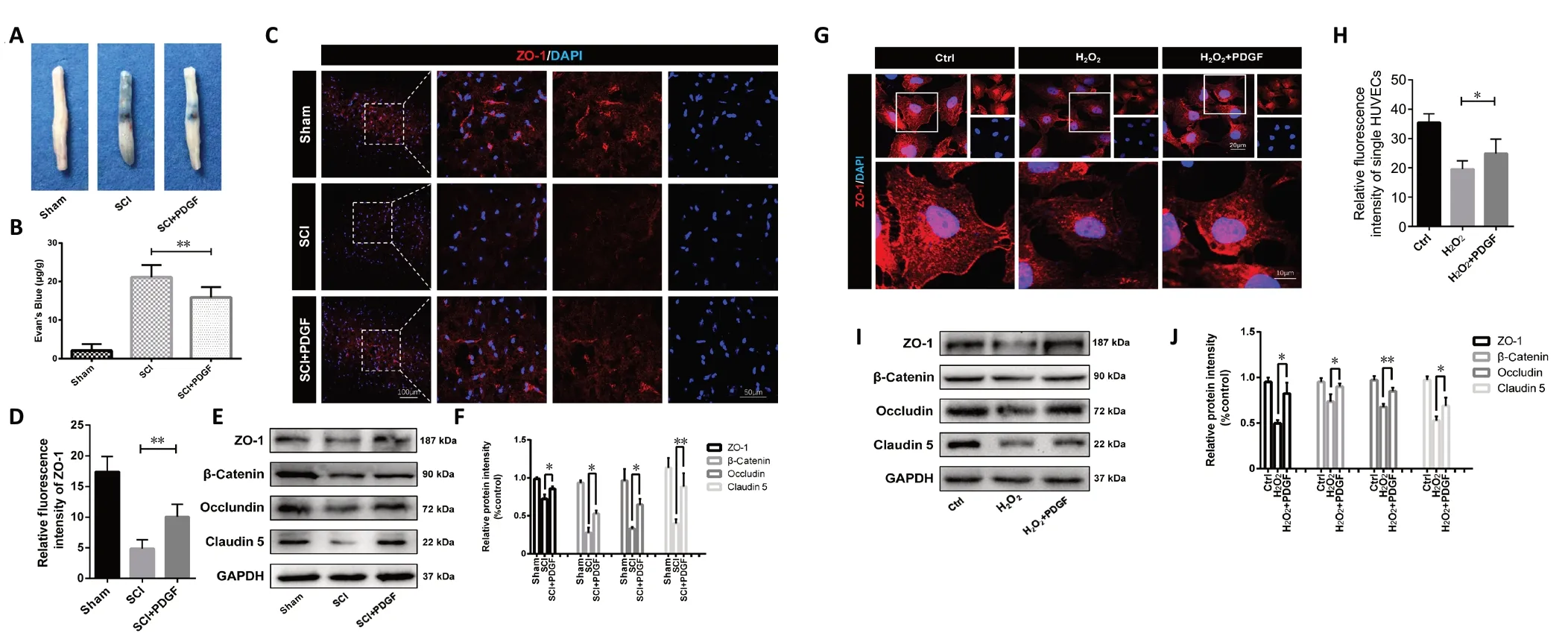
Figure 2 |Platelet-derived growth factor (PDGF) prevents blood-spinal cord barrier (BSCB) disruption by upregulating junction protein expression at 3 days post-spinal cord injury (SCI).
PDGF treatment relieves SCI-induced autophagic flux damage via the PDGFR/AKT signaling pathway
To determine whether PDGF treatment could decrease BSCB apoptosis after SCI through the PDGFR/AKT and autophagy signaling pathways, we next examined autophagy-related proteins in the damage areain vivo. As shown in Figure 4A–C, PDGF treatment activated phosphorylated PDGFR and AKT after SCI. Impaired autophagic flux has been shown to promote the pathogenesis of neurologic disease (Vidal et al.,2014). As shown in Figure 4K–L, autophagy protein (LC3 and p62) expression was up-regulated in the SCI group, indicating that autophagic flux was reduced after SCI, and the effect was markedly suppressed by PDGF treatment. Moreover,apoptosis-related proteins were also inhibited by PDGF (Figure 4M–O). We examined autophagy-related proteinsin vivoandin vitroto explore the relationship between autophagy and BSCB recovery. The number of LC3-positive cells was noticeably augmented in vessels of in the SCI group, while PDGF reversed this trend (Figure 4I–J). Consistent with thein vivoresults, cell experiments also confirmed that PDGF could inhibit the blocking of H2O2-induced autophagy in human umbilical vein endothelial cells (Figure 4D–H). These results revealed that relief of autophagic flux damage, especially in the BSCB, is involved in the protective effect of PDGF.
CQ reverses the protective effect of PDGF on BSCB integrity and function recovery
To investigate the effect of PDGF on BSCB integrity and function recovery, CQ (a blocking agent of autophagy) was used to evaluate the efficacy of PDGF post-SCI. As shown in Figure 5A–C, the increased expression of autophagy proteins(LC3 and p62) in the CQ group post-SCI suggests that CQ could inhibit the autophagy process. Next, we assessed the impact of autophagy on PDGF-mediated regulation of BSCB integrity and functional recovery. Compared with the SCI + PDGF group,AJ and TJ protein expression and apoptosis-related proteins were remarkedly inhibited in the PDGF with CQ treatment group (Figure 5D–H). Meanwhile, the results of Basso Mouse Scale scoring and the footprint analysis also indicated that CQ could reverse the protective effect of PDGF on functional recovery (Figure 5I and J). These findings suggested that the autophagy inhibitor CQ could eliminate the protective effect of PDGF on BSCB integrity and functional recovery post-SCI.

Figure 3 |Platelet-derived growth factor (PDGF) protects the neurovascular unit post-spinal cord injury (SCI).

Figure 4 |The protective effect of platelet-derived growth factor (PDGF) is associated with autophagic flux activated via the PDGFR/AKT signaling pathway after spinal cord injury (SCI).
Discussion
The BSCB is a tight barrier that maintains the normal functions of the spinal cord (Fassbender et al., 2011; Lee et al., 2014).The concept of NVU dysfunction has been described in stroke and other neurodegenerative diseases, and to date,an increasing number of studies have investigated the role of the NVU in SCI (Khan et al., 2009; Ahmad et al., 2012,2013). The link between NVU dysfunction and the BSCB in SCI is unclear. As a neurotrophic factor, PDGF has been used to treat Parkinson’s disease in clinical trials (Paul et al.,2015). Furthermore, PDGF has been reported to modulate endothelial proliferation and angiogenesis (Hellberg et al.,2010; Zhang et al., 2013a). Nevertheless, whether PDGF-BB can be used as potential treatment for SCI patients remains unclear. In this study, PDGF was found to promote recovery in SCI mice. Additionally, the protective function of PDGF post-SCI was correlated with the downregulation of autophagy, thereby ameliorating BSCB disorder and improving NVU recovery.
The BSCB is disrupted immediately after SCI, leading to a secondary injury cascade (Whetstone et al., 2003).Additionally, a previous study showed that basic FGF regulated junction proteins through the PI3K-AKT-Rac1 pathway to prevent blood-brain barrier injury post-TBI, revealing the potential function of growth factors in neuronal injury(Wang et al., 2016). Interestingly, PDGF is widely studied in neurological diseases such as Parkinson’s (Paul et al.,2015) and Alzheimer’s diseases (Masliah et al., 1995), while few studies have evaluated the effect of PDGF on SCI. The results showed that PDGF administration markedly facilitated functional recovery post-SCI, reduced Evans blue leakage,and protected the SCI area by increasing expression of junction proteins after SCI. Many studies have examined recovery of the BSCB; however, the mechanism by which BSCB repair affects functional recovery remains unclear. Notably,previous studies have shown that TBI can be prevented by reducing exacerbations of the NVU, which are associated with blood-brain barrier leakage induction, cell death, and neurobehavioral deficits in rats (Khan et al., 2009). Thus,expression levels of Lectin, NeuN, GFAP, and the extracellular matrix were examined, which showed that the number of neurons was augmented with increasing blood vessels, while GFAP-positive cells were reduced after 14 days. PDGF also significantly enhanced collagen expression. These results are consistent with those reported by Khan et al. (2009). Sweeney et al. (2016) noted that pericytes can regulate blood-brain barrier permeability, angiogenesis, and neuroinflammation,and the PDGF/PDGFRβ signaling pathway plays a crucial role in the proliferation, migration, and recruitment of pericytes in the vessel wall. Interestingly, Oztop-Cakmak et al. (2017)reported that, as a functionally vital component of the bloodbrain barrier, pericytes provide adequate microcirculatory supply and couple neurovascular function. We also found that PDGF could promote pericyte proliferation (Additional Figure 1). In our future studies, we aim to focus on the role of pericytes in NVU recovery post-SCI. Taken together, our results demonstrated for first time that the protective effect of PDGF on SCI might be related to BSCB repair and NVU recovery.
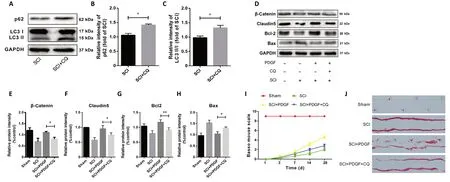
Figure 5 |Chloroquine (CQ) reverses the protective effect of platelet-derived growth factor (PDGF) on blood-spinal cord barrier integrity and functional recovery.
Alirezaei et al. (2011) found that the autophagy signal may directly facilitate cell death in neuronal injury-inducing diseases. Meanwhile, the autophagy process has been shown to be blocked during SCI progression (Wang et al., 2014).Moreover, Zhang et al. (2013b) suggested that basic FGF inhibits the autophagic protein LC3II and augments survival and recovery of neurons in a rat spinal cord model. Our study found that PDGF could activate autophagic flux in endothelial cells. Wang et al. (2012) suggested that AKT regulates autophagy and tumorigenesis via Beclin1 phosphorylation.Zhang et al. (2019) suggested that autophagy induced by oxygen-glucose deprivation mediates NVU injury. In addition,Wu et al. (2020) demonstrated that Dl-3n-butylphthalide inhibits autophagy-induced blood-brain barrier disruption and cell apoptosis. These results suggest a close relationship between autophagy and the NVU, which requires further research. In this study, the PDGF group activated PDGFR/AKT signaling after SCI, indicating that PDGF might enhance TJ and AJ protein expression by the PI3K/AKT signaling pathway to activate autophagic flux.
The critical function of autophagy in both TBI and SCI is controversial, particularly regarding the BSCB. Therefore,it is not sufficient to explain the role of autophagy in SCI by activation or inhibition alone; further investigation of the different types of cells or different time points in SCI is required (Tang et al., 2014). In this study, CQ (a blocking agent of autophagy) abolished the protective effect of PDGF on BSCB integrity and function recovery, implying that autophagic flux activity is involved in the protective function of PDGF.Overall, these findings indicated that the protective effect of PDGF on the BSCB might be associated with autophagic flux activity and may improve NVU recovery in SCI models.
In conclusion, we confirm that PDGF protects against BSCB disorders and neuron apoptosis, thereby promoting motor function recovery. Additionally, our findings offer a new perspective in that repair of the BSCB induces recovery of the NVU, thus enhancing functional recovery after SCI. PDGF treatment triggers the PDGFR/AKT and autophagy pathway.Overall, PDGF may comprise a potential intervention to suppress BSCB disruption and promote recovery after SCI.
Author contributions:Study conception and design: LXY, NCA, PH; data acquisition: DHL, ZLZ, HJ, HL; data analysis and interpretation: DQC, YQW, JX;manuscript writing: KX, XKL; manuscript review: HYZ. All authors approved the final version of the manuscript.
Conflicts of interest:The authors declare no competing interests.
Financial support:This study was partly supported by research grants from the National Natural Science Foundation of China, Nos. 81802251 (to KX),81772450 (to HYZ) and 81801233 (to YQW); the Natural Science Foundation of Zhejiang Province of China, Nos. LQ18H150003 (to KX), LY19H150001(to DQC), LQ18H090011 (to YQW) and LQ20C200015 (to HJ); and the Opening Project of Zhejiang Provincial Top Key Discipline of Pharmaceutical Sciences, No. YKFJ3-011 (to KX). The funders had no roles in the study design,conduction of experiment, data collection and analysis, decision to publish, or preparation of the manuscript.
Institutional review board statement:The study was approved by the Ethics Committee of Wenzhou Medical University (approval No. wydw2018-0043) in July 2018.
Copyright license agreement:The Copyright License Agreement has been signed by all authors before publication.
Data sharing statement:Datasets analyzed during the current study are available from the corresponding author on reasonable request.
Plagiarism check:Checked twice by iThenticate.
Peer review:Externally peer reviewed.
Open access statement:This is an open access journal, and articles are distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 License, which allows others to remix, tweak,and build upon the work non-commercially, as long as appropriate credit is given and the new creations are licensed under the identical terms.
Open peer reviewers:Shinichi Takahashi, Keio University School of Medicine,Japan; Miao Zhang, Harvard School of Dental Medicine, USA.
Additional files:
Additional Figure 1:Growth-promoting effects of PDGF in pecitytes.
Additional file 1:Open peer review reports 1 and 2.

- 中国神经再生研究(英文版)的其它文章
- The use of hydrogel-delivered extracellular vesicles in recovery of motor function in stroke: a testable experimental hypothesis for clinical translation including behavioral and neuroimaging assessment approaches
- Advances in human stem cell therapies: pre-clinical studies and the outlook for central nervous system regeneration
- MicroRNAs in laser-induced choroidal neovascularization in mice and rats: their expression and potential therapeutic targets
- The emerging role of probiotics in neurodegenerative diseases: new hope for Parkinson’s disease?
- The phenotypic convergence between microglia and peripheral macrophages during development and neuroinflammation paves the way for new therapeutic perspectives
- Modeling subcortical ischemic white matter injury in rodents: unmet need for a breakthrough in translational research