
玉米跨代干旱胁迫记忆生理机制及DNA甲基化变化分析
2021-11-09 11:43尹晓雨于晓明刘晓丹
西北植物学报 2021年10期
王 霞,尹晓雨,于晓明,刘晓丹
(吉林农业科技学院,吉林 吉林 132101)
玉米(ZeamaysL.)不仅是中国三大粮食作物之一,还是重要的饲料作物和工业原料,是目前世界上产量最大的禾谷类作物,在国民经济中占有非常重要的地位。作为旱地农作物,玉米的需水量非常大,对环境中的水分缺失很敏感[1]。随着全球干旱问题日益严重,干旱已经成为制约玉米产量的首要因素,中国约有60%的玉米种植面积受到干旱胁迫[1]。作为中国玉米主产区,吉林省西部有一半以上的干旱地没有得到开发利用。研究玉米的抗旱机制、培育抗旱新品种是目前亟待解决的问题,而弄清玉米跨代干旱胁迫记忆机制是解决这一问题的重要途径。
胁迫记忆(stress imprint)是指植物在经历初始胁迫后,当再次面对生物和非生物胁迫(干旱、冷冻、高温、高盐等)时会产生更高耐受性的现象[2]。2015年,Ramírez[3]等发现经多次干旱胁迫后的马铃薯拥有更强的抗氧化性,同样,研究者们在小麦[4]、葡萄[5]、马铃薯[5]等多种植物中陆续发现它们对过去经历的逆境胁迫具有记忆效应。2018年,“植物对逆境的记忆功能与进化”入选中国科学技术协会的60个重大科学问题和工程技术难题,掀起了胁迫记忆相关研究的热潮。研究发现,胁迫记忆可能是短期的,也可能是长期的,短期记忆不能遗传,但可以提高植物对逆境的适应能力,长期记忆却可以通过一种未知的机制传递给子代,产生跨代胁迫记忆,使植物能够更快更强地应对逆境[3]。目前,大量证据表明,植物会对当代胁迫产生印记并表现出更为强烈的抵御机制[3-6],但是关于跨代胁迫记忆方面的研究报道却不多。2009年,Whittle等[7]发现F0及F1代拟南芥经高温胁迫后,其F3代仍具有较强的抵抗高温的能力;2015年,Shi等[8]发现水稻经高温胁迫锻炼后,其子代对高温的耐受性明显增强;2019年,Cong等[9]的研究表明,由重金属胁迫引起的水稻HMA基因表达上调产生了跨代胁迫记忆,这种现象同样也在马铃薯[3]、拟南芥[10]中被发现。
迄今,植物胁迫记忆产生与维持的具体机制尚不清楚。现有研究表明,胁迫记忆的产生和维持与胁迫诱导的代谢产物、DNA甲基化及组蛋白修饰等有关[11]。作为最重要的表观遗传修饰之一,DNA甲基化修饰在植物非生物胁迫应答中的作用引起了研究者们的极大兴趣。2016年,Wibowo等[12]发现,盐胁迫可以引起拟南芥产生跨代胁迫记忆,且这种记忆的产生与维持和特定DNA的超甲基化有关,且当盐胁迫消失时,DNA的超甲基化也会逐渐消失;2019年,Li等[13]发现水稻经多次干旱胁迫后,约3 000多个记忆转录本与DNA甲基化调控有关。对胁迫记忆中DNA甲基化变化机制的深入研究,可以为我们从表观遗传修饰层面改良植物抗逆性奠定基础。目前,针对玉米跨代胁迫记忆的研究尚无报道,本试验以玉米模式自交系B73和H99为研究对象,通过分析2个玉米品种G0和G1代响应干旱胁迫时生理指标和基因组DNA甲基化修饰的变异和传递规律,为后续跨代胁迫记忆机制研究奠定基础。
1 材料和方法
1.1 试验材料及处理
1.1.1 试验材料参试玉米自交系B73和H99均为引自美国的玉米经典自交品系,两者均较早完成基因组测序,都为玉米育种的种质基础,试验种子由吉林省农业科学院提供。
1.1.2 干旱锻炼参试材料为B73和H99在正常水分条件下套袋授粉获得单株种子,采用纸皿法发芽,待胚芽破胸露白时,每个品种挑选300粒发芽良好的种子,进行干旱锻炼。每天上午9:00将发芽种子置于干滤纸上,于22 ℃光照培养箱(光照强度400 μmol·m-2·s-1、湿度60%)中干旱锻炼1 h,然后置于湿纱布中,每天锻炼1次,连续锻炼4 d(1 h→2 h→3 h→4 h),累计锻炼4 h,对照组(0 h)不进行干旱锻炼(图1,A),之后2类经过处理的发芽种子均栽种于吉林农业科技学院玉米试验田,经套袋授粉后分别获得子代种子,分别记为G0(对照)和G1(锻炼)代。
1.1.3 干旱胁迫处理G0和G1代玉米种子采用纸皿法催芽后种植,待幼苗长至五叶一心时,取出幼苗,洗净根部泥沙,将幼苗根部放入1/2MS液体培养基中适应2 d,然后移入含20% PEG-6000的1/2MS液体培养基中,分别在浸泡0、1、2、3、4、5 h后(图1,B),随机取16棵玉米幼苗第四叶混样,保存于-80 ℃中备用。
1.2 测定指标及方法
1.2.1 生理指标采用饱和称重法测定叶片相对含水量(relative water content, RWC)[6],硫代巴比妥酸法测定丙二醛(malondialdehyde, MDA)含量[14],蒽酮法测定可溶性糖含量[15],茚三酮显色法测定脯氨酸含量[15],氮蓝四唑(NBT)法测定超氧化物歧化酶(superoxide dismutase,SOD)活性[15],愈创木酚法测定过氧化物酶(peroxidase,POD)活性[14],设3个重复。
1.2.2 DNA甲基化敏感扩增多态性(MSAP)采用改良CTAB法提取玉米叶片基因组DNA,参照孙丽芳等[16]的方法进行MSAP分析,设3个重复。参照Zhang等[17]的方法,统计并分析DNA甲基化水平和模式变异情况,EcoRⅠ、HpaⅡ和MspⅠ购自大连宝生物,接头及引物均由上海生工公司合成,序列见表1。

图1 玉米自交系B73在干旱锻炼(A)与干旱胁迫(B)后的表型变化Fig.1 Phenotypic changes of maize self-bred line B73 after drought hardening (A) and drought stress (B)
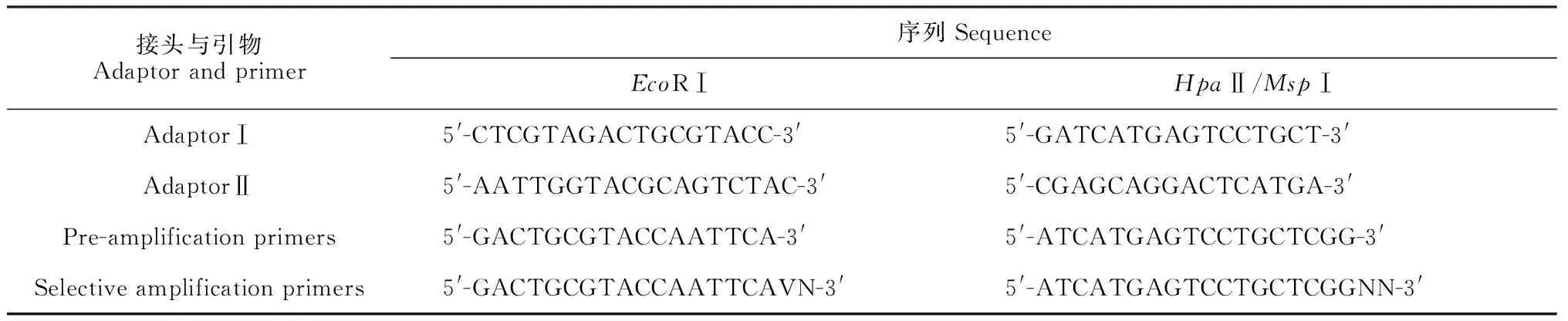
表1 MSAP用的接头及引物序列
1.3 抗旱性综合评价
依据生理指标数据采用隶属函数法[18]进行玉米幼苗抗旱性综合评价,公式为:
指标与抗性正相关:
X(u) = (X-Xmin) /(Xmax-Xmin);
指标与抗性负相关:
X(u) =1- (X-Xmin) /(Xmax-Xmin)。
其中,X为指标平均值,Xmax、Xmin分别为指标最大值和最小值。
1.4 数据分析
用Excel 2010作图,单因素方差分析与显著性检验(Duncan法,P<0.05)用SPSS26.0软件。
2 结果与分析
2.1 干旱胁迫对G0和G1代玉米幼苗叶片相对含水量(RWC)的影响
由图2,A可知,所有材料叶片RWC均随胁迫时间延长呈逐渐下降趋势,且各干旱胁迫处理时期均存在显著性差异(P<0.05)。同一玉米品种内相比较,G1代叶片RWC在未经胁迫时稍高于G0代(P>0.05),在经过相同时间干旱胁迫后均显著高于G0代(P<0.05);在胁迫5 h时,B73和H99玉米 G1代叶片RWC分别是G0代的1.14和1.09倍,且2个品种G1代叶片RWC的降幅均小于G0代。在品种间相比较,经过相同时间干旱胁迫后,B73自交系 G1和G0代叶片RWC均高于H99,它们叶片RWC降幅也均显著低于H99自交系。
2.2 干旱胁迫对G0和G1代玉米幼苗叶片MDA含量的影响
干旱胁迫可显著提高两个玉米自交系叶片MDA含量(图2,B)。随干旱胁迫时间延长,所有材料叶片MDA含量均呈显著上升趋势(P<0.05);对于同一玉米自交系,其G1代叶片MDA含量在未经胁迫时稍低于G0代(P>0.05),在经过相同时间胁迫后均显著低于G0代(P<0.05);在胁迫5 h时,B73和H99 自交系G1代叶片MDA含量分别是G0代的0.85和0.90倍,且2个自交系G1代叶片MDA含量增幅均小于G0代。在品种间相比较,经过相同时间干旱胁迫后,B73 自交系G1和G0代叶片MDA含量均显著低于H99自交系(P<0.05)。
2.3 干旱胁迫对G0和G1代玉米幼苗叶片对可溶性糖和脯氨酸含量的影响
干旱胁迫可显著提高玉米自交系叶片可溶性糖含量,且所有材料叶片可溶性糖含量均随着胁迫时间延长呈上升趋势(图2,C)。除未经胁迫时,H99玉米自交系G1代叶片可溶性糖含量稍高于G0代(P>0.05)外,同一玉米自交系经过相同时间胁迫后G1代叶片可溶性糖含量均显著高于G0代(P<0.05);在干旱胁迫5 h时,B73和H99 自交系G1代叶片可溶性糖含量分别是G0代叶片的1.17和1.11倍,且2个自交系G1代叶片可溶性糖增加量高于G0代。经过相同干旱胁迫后,B73 玉米自交系G1和G0代叶片可溶性糖含量均显著高于H99自交系(P<0.05)。

不同字母表示处理材料及时间之间在0.05水平差异显著(P<0.05),下同图2 B73和H99玉米自交系G0、G1代幼苗叶片4个生理指标比较Different normal letters indicate significant differences among materials and times at 0.05 level(P<0.05), the same as belowFig.2 Comparison of four main indexes in leaves of G0 and G1 generation seedlings from maize self-bred line B73 and H99
同时,图2,D显示,干旱胁迫可提高玉米自交系叶片脯氨酸含量,且所有材料叶片脯氨酸含量均随胁迫时间延长呈逐渐上升趋势。对于同一玉米自交系,G1代叶片脯氨酸含量在未经干旱胁迫时稍高于G0代(P>0.05),在经过相同时间胁迫后均显著高于G0代(P<0.05);在干旱胁迫5h时,B73和H99 玉米自交系G1代叶片脯氨酸含量分别是G0代的1.23和1.16倍,且2个自交系G1代叶片脯氨酸增加量也均高于G0代。自交系间相比较,经过相同时间干旱胁迫后,B73 玉米自交系G1和G0代叶片脯氨酸含量均显著高于H99自交系(P<0.05)。
2.4 干旱胁迫对G0和G1代玉米幼苗叶片抗氧化酶活性的影响
2.4.1 SOD活性的影响由图3,A可知,随干旱胁迫时间延长,所有材料叶片SOD活性均呈先升后降趋势,并均在胁迫4 h时达到最高,此时B73和H99 自交系G1代叶片SOD活性分别是G0代的1.04和1.02倍;就同一玉米自交系而言,G1代叶片SOD活性在未经干旱胁迫时稍高于G0代(P>0.05),在经过相同时间干旱胁迫后均显著高于G0代(P<0.05),且G1代叶片SOD活性增加量均高于G0代。经过相同时间干旱胁迫后,B73 自交系G1和G0代叶片SOD活性均显著低于H99自交系(P<0.05),但是B73 叶片G1和G0代SOD活性增加量却明显高于H99自交系。
2.4.2 POD活性图3,B显示,随干旱胁迫时间延长,所有材料叶片POD活性均呈先升后降趋势,并均在胁迫3 h时达到最高值,此时B73和H99 自交系G1代叶片POD活性分别是G0代的1.10和1.06倍;在同一玉米自交系内,G1代叶片POD活性在未经干旱胁迫处理时稍高于G0代(P>0.05),在干旱胁迫处理1 h时,H99自交系 G1代叶片POD活性稍高于G0代(P>0.05),其余处理组G1代POD活性均显著大于G0代(P<0.05),且G1代叶片POD活性增加量也均高于G0代。另外,经过相同时间干旱胁迫后,B73自交系 G1和G0代叶片POD活性均显著低于相应的H99自交系(P<0.05),而B73 自交系G1和G0代叶片POD活性增加量却明显高于相应的H99自交系。
2.5 干旱胁迫对G0和G1代玉米幼苗叶片DNA甲基化修饰的影响
从48对引物中筛选出20对高多态性引物进行PCR扩增,B73和H99玉米自交系的扩增总条带数分别为632和589,平均每对引物扩增条带数分别为31.60和29.45,2个玉米自交系经干旱胁迫后,同一自交系相同世代DNA甲基化水平及模式变化均不明显,故实验只统计干旱胁迫0、1、3、5h的甲基化状况。
2.5.1 DNA甲基化水平由图4可知,随干旱胁迫时间延长,B73 自交系2个世代(G0和G1代)CG、CHG甲基化水平,以及H99 自交系2个世代CG甲基化水平均呈缓慢上升趋势,H99 自交系2个世代CHG甲基化水平均呈缓慢下降趋势,但其变化均未达到显著水平(P>0.05);干旱胁迫期间,B73 自交系G1代CG+CHG甲基化水平(平均值30.30%)略高于G0代(平均值30.21%),H99 自交系G1代CG+CHG甲基化水平(平均值29.96%)略低于G0代(平均值30.10%),B73和H99 自交系G1代CG、CHG甲基化水平变化量均大于G0代,但其变化均未达到显著水平(P>0.05)。

图3 B73和H99玉米自交系G0、G1代叶片抗氧化酶活性比较Fig.3 Comparison of antioxidant enzyme activities between G0 and G1 generation seedlings from maize self-bred line B73 and H99
2.5.2 DNA甲基化模式植物基因组DNA甲基化主要以胞嘧啶甲基化为主,其模式有CG超甲基化(CG hyper)、 CG去甲基化(CG hypo)、 CHG超甲基化(CHG hyper)、CHG去甲基化(CHG hypo)、both hyper和both hypo 6种。由图5可知,干旱胁迫可引起B73和H99 玉米自交系2个世代DNA甲基化模式发生显著变化(P<0.05);随干旱胁迫时间延长,所有材料6种DNA甲基化模式的变化均缓慢增强,G1代DNA甲基化模式变化均稍高于G0代;B73 自交系2个世代DNA甲基化模式均以CG hypo和CHG hypo变化为主,其次是CHG hyper和CG hyper变化,而H99 自交系2个世代均以CHG hypo和CG hyper变化为主,其次是CG hypo和CHG hyper变化;2个自交系G1和G0代的DNA甲基化模式both hypo与both hyper 变异率均极低。其中,经20%PEG干旱胁迫5 h,B73 自交系G1和G0代的甲基化模式总变异率分别为3.090%和0.208%,H99 自交系G1和G0代的甲基化模式总变异率分别为3.562%和0.223%,B73 自交系2个世代整体甲基化模式变异率小于H99自交系。

图4 B73和H99玉米自交系G0、G1代叶片基因组DNA甲基化水平比较Fig.4 Comparison of DNA methylation levels of G0 and G1 generation seedlings from maize self-bred line B73 and H99
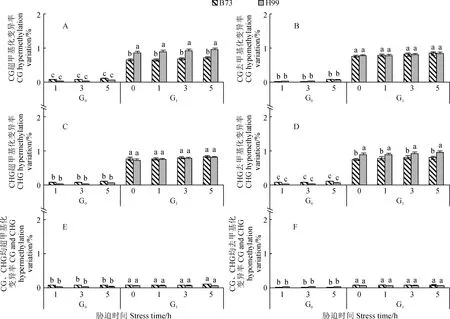
图5 B73和H99玉米自交系G0、G1代叶片基因组DNA甲基化模式比较Fig.5 Comparison of DNA methylation patterns of G0 and G1 generation seedlings from maize self-bred line B73 and H99
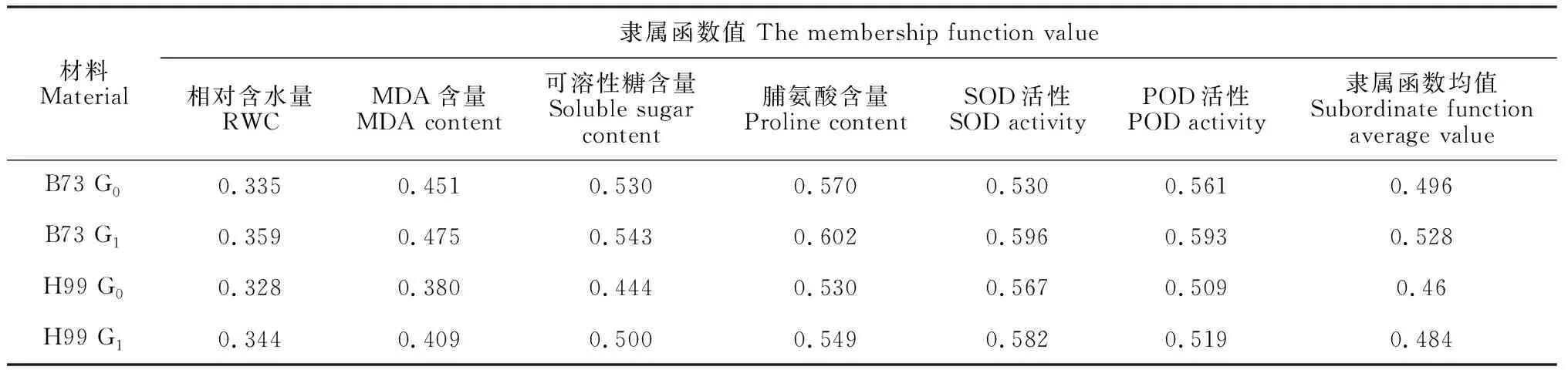
表2 4份玉米材料的抗旱隶属函数值
2.6 玉米自交系G0和G1代幼苗抗旱性综合评价
目前尚无植物抗旱性评价的标准指标,干旱环境下植物的生理变化非常复杂,利用单一生理指标评价抗旱性并不科学,隶属函数法是以各生理指标隶属度的平均值作为标准来评价植物的抗旱性,平均值越高,抗旱性越强[18]。两个玉米自交系G0和G1代6个指标的隶属函数计算结果(表2)显示,4份材料的抗旱性由强到弱依次为:B73 G1代>B73 G0代>H99 G1代>H99 G0代,说明B73和H99自交系经干旱锻炼后,其自交后代的抗旱性均有所增强,且B73抗旱性高于H99。
3 讨 论
3.1 干旱锻炼对玉米自交系G1代响应干旱胁迫时生理指标的影响
已有的干旱胁迫经历可能会改变植物应对干旱胁迫的生理反应。RWC是直接评价植物保水能力的指标,RWC值越小,说明植物缺水越严重,受干旱胁迫损伤程度越深[19],因此RWC常被用于评价植物的抗旱性。Ding等研究发现,经多次短时干旱锻炼后,玉米[6]和拟南芥[20]当代叶片RWC均明显提高。本实验中,在干旱胁迫条件下,B73和H99 玉米自交系G1代叶片RWC均显著高于G0代,说明G0代干旱锻炼提高了G1代叶片的保水能力。可以推测,一定强度的干旱锻炼可以提高玉米植株当代、甚至其子代叶片的保水能力。同时,B73和H99 玉米自交系G1和G0代叶片RWC随干旱胁迫强度增加显著下降,具有一定的剂量效应。
缺水可以引起植物细胞内活性氧的大量累积[21],引发膜脂过氧化反应,该反应的最终产物MDA会破坏膜的结构和功能,甚至导致植物死亡。因此,MDA含量是衡量植物受干旱伤害程度的重要指标[14]。与此同时,多数植物还拥有一套清除活性氧的抗氧化保护系统,其主要成员有SOD和POD等抗氧化酶,干旱条件下植物可以通过增加这些抗氧化酶的活性来避免或减轻干旱带来的伤害[21]。本实验结果表明,干旱胁迫可以引起B73和H99玉米自交系叶片的膜损伤,改变其抗氧化酶活性,同时,与G0代相比,B73和H99 自交系G1代经历干旱胁迫时的MDA含量增加速度均明显减缓,SOD和POD活性均明显提高,说明G0代玉米干旱锻炼可提高G1代的抗氧化能力,降低由干旱胁迫带来的膜伤害,这与拟南芥[17]、马铃薯[5]、小麦[18]的相关研究结果一致。另外,本实验中,当干旱胁迫超过4 h后,所有材料叶片的SOD和POD活性均开始下降,证明高强度干旱胁迫会破坏植物的自身抵御能力[22]。
渗透调节机制同样是植物适应干旱的重要途径,植物借助脯氨酸、可溶性糖等渗透调节物质使细胞维持一定的膨压,使膜蛋白维持正常结构、参与活性氧的清除[21]。因此,研究者们认为由逆境引起的渗透调节物质的累积与植物的抗旱性成正相关[14]。本实验恰好证明了这一结论:B73和H99 自交系G1代再次经历干旱胁迫时,其叶片脯氨酸和可溶性糖含量均明显高于G0代,即亲代干旱锻炼可增加玉米自交后代渗透调节物质的累积,降低细胞渗透势,利于细胞保水,这与在小麦[18]上相关的研究结果一致。
3.2 干旱锻炼对玉米自交系G1代响应干旱胁迫时DNA甲基化修饰的影响
DNA甲基化修饰不仅参与植物对逆境胁迫的应答,还参与植物抗逆胁迫记忆的形成与维持,现已在水稻[23]、番茄[24]、拟南芥[25]中发现了与DNA甲基化修饰相关的基因参与了抗逆应答; Wibowo等[12]则发现水稻跨代盐胁迫记忆与特定区域DNA超甲基化有关;Herman与Boyko则分别证明DNA甲基化介导了桃叶寥[26]和拟南芥[10]的跨代干旱胁迫记忆。本实验中,同一玉米自交系2个世代DNA甲基化水平变化趋势相同,但是G1代DNA甲基化修饰变化大于G0代,与水稻[27]的相关研究结果吻合。在干旱胁迫下,B73和H99 玉米自交系2个世代基因组DNA甲基化水平和甲基化模式的变化并不明显,推测DNA甲基化修饰的改变可能需要相对较长时间,同时B73和H99甲基化修饰响应胁迫的变异规律并不相同,证明了Ferreira提出的DNA甲基化动态变化与基因型有关的说法[28]。
与G0代相比,2个玉米自交系G1代叶片CG、CHG甲基化修饰均更不稳定,G1代抗旱性均有所增强,说明G0代的干旱锻炼在G1代体内留有印记,当再次遭受干旱逆境时,可以产生更快更强的DNA甲基化变化,启动自身防御系统,提升保水能力,避免或减缓伤害,进而表现出更强的抗旱性。本研究从生理和DNA甲基化修饰视角分析了干旱锻炼对玉米自交后代抗旱性的影响,为玉米抗旱新品种培育及植物跨代胁迫记忆机制研究奠定了基础。
猜你喜欢
北京农学院学报(2019年1期)2019-02-22
浙江农业学报(2017年1期)2017-05-17
河南农业科学(2017年4期)2017-04-12
西南农业学报(2016年5期)2016-05-17
西南农业学报(2016年4期)2016-05-17
西南农业学报(2016年6期)2016-04-16
山西农经(2015年7期)2015-07-10
现代检验医学杂志(2015年2期)2015-02-06
沈阳医学院学报(2014年4期)2014-12-27
遗传(2014年3期)2014-02-28
