
安徽省不同地区日本血吸虫群体线粒体基因遗传变异研究
2021-11-10 03:29马晓荷汪敏朱磊郭见多李清越刘婷翟杜娟孙成松张世清汪天平
热带病与寄生虫学 2021年5期
马晓荷,汪敏,朱磊,郭见多,李清越,刘婷,翟杜娟,孙成松,张世清,汪天平
安徽省血吸虫病防治研究所,安徽 合肥 230601
血吸虫病是由裂体吸虫属吸虫感染引起的寄生虫病,该病严重危害人类健康,阻碍经济社会发展,主要分布在非洲、亚洲和拉丁美洲,是全球重要的公共卫生问题[1-2]。感染人类的血吸虫之中,中国只存在日本血吸虫[1,3]。安徽省日本血吸虫病流行区类型以湖沼型为主[4-6],还包括洲滩型和山丘型。
在地理隔离[7-8]、终宿主差异[9]等因素的影响下,血吸虫个体会产生遗传变异。线粒体DNA(mtDNA)是寄生虫群体遗传研究中具有较好价值的遗传标志[10]。单个个体内mtDNA有高度均一性,遵循严格的母系遗传方式,具有进化速度快、群体内变异大、近缘种间解释的灵敏度高等特点,利于进行群体遗传分析。为了解安徽省不同地区日本血吸虫群体间的遗传进化关系,本文通过测定并分析安徽省不同类型流行区日本血吸虫成虫的线粒体NADH(还原型烟酰胺腺嘌呤二核苷酸)脱氢酶1(ND1)和细胞色素C氧化酶亚基Ⅰ(COⅠ)基因序列并构建系统进化树,探讨安徽省不同类型血吸虫病流行区日本血吸虫群体间的进化关系。
1 材料与方法
1.1材料 采集安徽省山丘型流行区(池州市石台县)和湖沼型流行区(安庆市大观区、铜陵市枞阳县)的钉螺,逸蚴筛选阳性钉螺并继续饲养。两周后对钉螺进行再次逸蚴,收集尾蚴感染BALB/c小鼠(雌性,6~7周),35天后解剖小鼠,门静脉灌注法分离和收集成虫,单只成虫分别收集于含有95%乙醇的1.5 mL离心管中,-80 ℃保存。
1.2方法
1.2.1日本血吸虫成虫DNA提取 每只成虫用pH为7.4的磷酸盐缓冲液(NaCl 137 mmol/L,KCl 2.7 mmol/L,Na2HPO44.3 mmol/L,KH2PO41.4 mmol/L)洗涤3次,去除杂质和95%乙醇。用DNeasy Blood & Tissue Kit试剂盒(Qiagen,德国)提取DNA,具体操作流程参照说明书,不同地区的血吸虫成虫DNA单独提取。
1.2.2ND1和COⅠ基因PCR扩增和测序 以GenBank中日本血吸虫线粒体基因组序列NC_002544.1中COⅠ基因编码序列和ND1基因编码序列为模板设计引物。参考序列COⅠ基因长度为1 527 bp,正向引物5'-GATTTGATATCTTGATTTGTCT-3',反向引物5'-CTAATACTTTGATCAATTCTTACCG-3';ND1基因长度为891 bp,正向引物5'-ATGAATATATGAAACTTTTTAG-3',反向引物5'-CTACCTTATAATTATAATCAATCCG-3'。PCR扩增反应体系总体积50 μL,ND1扩增条件:94 ℃预变性5 min,94 ℃ 30 s,50 ℃ 30 s,72 ℃ 1 min,35个循环,72 ℃延伸8 min;COⅠ扩增条件:94 ℃预变性5 min,94 ℃ 30 s,50 ℃ 30 s,72 ℃ 90 s,35个循环,72 ℃延伸8 min。1%琼脂糖凝胶电泳检测PCR扩增产物。
1.2.3ND1和COⅠ基因克隆和测序 用胶回收试剂盒纯化扩增产物,然后将产物连接至pESI-T载体(上海翊圣生物科技有限公司),转化大肠杆菌DH5α感受态细胞,筛选重组阳性克隆。将鉴定成功的克隆子送至上海生工生物工程有限公司测序。
1.2.4序列分析及构建系统进化树 利用Chromas软件查看测序峰图,手工筛查每条序列的测序结果。利用Clustal X 2.0[11]比对序列,使用DNAsp 6.0[12]和Mega X[13]分析序列的基本特征。在Mega X中以COⅠ和ND1序列构建系统进化树,外群(outgroup)分别为曼氏血吸虫(Schistosomamansoni)的COⅠ和ND1序列,序列来源为GenBank中曼氏血吸虫线粒体全基因组序列,序列号为NC_002545.1。建树方法为邻接法(Neighbor-Joining Method),自展值(bootstrap)为1 000,样本间进化距离计算方法为Kimura 2-parameter 算法。
2 结 果
2.1ND1和COⅠ基因序列多态性分析 本研究中所有样品扩增获得的条带大小均与预期一致,截去首尾部分序列后用于分析的ND1和COⅠ基因序列长度分别为844 bp和1 481 bp。ND1基因序列存在15个变异位点,G+C含量为0.298,碱基转换数为3,没有发生碱基颠换;COⅠ序列存在74个变异位点,G+C含量为0.326,碱基转换数为21,碱基颠换数为5。与ND1基因相比,COⅠ基因序列的突变位点较多,碱基替换种类中不仅存在碱基转换还存在碱基颠换。样本的COⅠ序列核苷酸多样性(Pi)为0.017 3,高于ND1(0.004 7),综合其他数据可见所研究地区日本血吸虫的COⅠ基因比ND1基因具有更高的序列多样性。见表1。
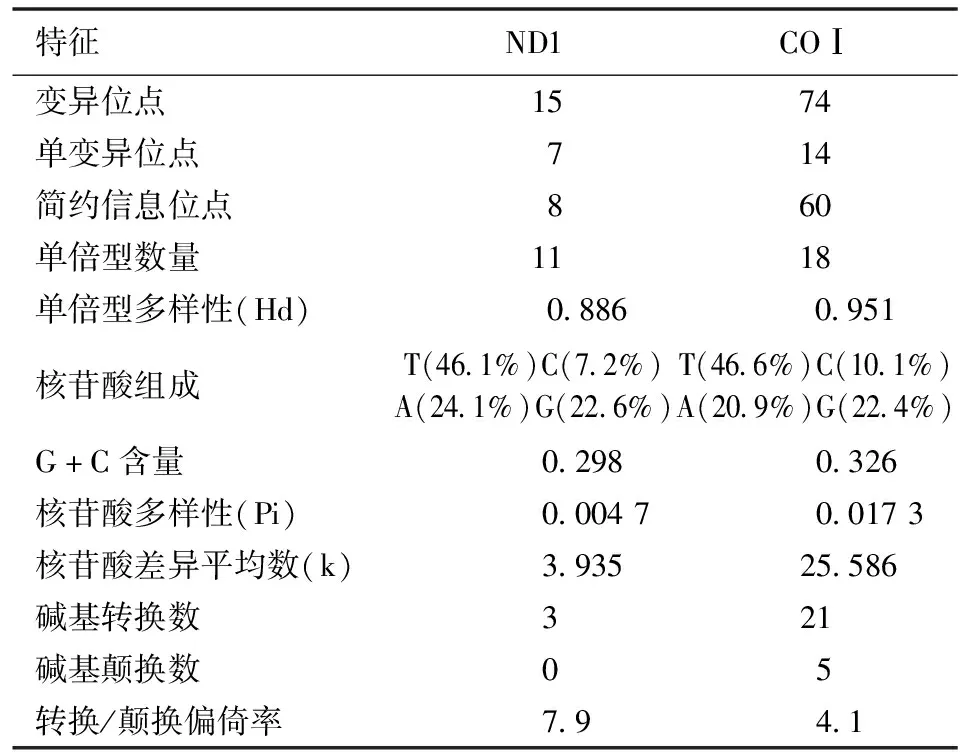
表1 安徽省日本血吸虫线粒体ND1和COⅠ基因序列的基本特征
对不同地区血吸虫序列进行比较发现,来自湖沼型流行区的日本血吸虫样本ND1的基因变异位点数、核苷酸多样性(Pi)、单倍型个数和单倍型多样性(Haplotype diversity,Hd)均高于来自山丘型流行区的样本。来自湖沼型流行区的日本血吸虫样本COⅠ序列的变异位点数、核苷酸多样性也明显高于来自山丘型流行区的样本。见表2。

表2 安徽省不同地区日本血吸虫线粒体ND1和COⅠ基因序列比较
2.2不同地区日本血吸虫种群间遗传距离和进化分析 不同地区日本血吸虫成虫的ND1和COⅠ基因序列均存在略微差异。来自山丘型流行区与两处来自湖沼型流行区的日本血吸虫样本之间,ND1序列遗传距离分别为0.004 8和0.003 3;COⅠ序列遗传距离均为0.035。两处来自湖沼型流行区的日本血吸虫样本之间ND1和COⅠ序列遗传距离为0.001 8和0.007。
依据COⅠ 和ND1序列构建的系统进化树显示,来自山丘型流行区和湖沼型流行区的日本血吸虫样本ND1序列没有较大的差异,见图1A。而COⅠ序列构建的系统进化树中,来自山丘型流行区的日本血吸虫样本单独成簇,且Bootstrap值为100%,说明和来自湖沼型流行区的日本血吸虫样本相比,来自山丘型流行区的日本血吸虫样本COⅠ基因序列发生了一定程度的变化,见图1B。

注:A为基于ND1序列的系统进化树,B为基于COⅠ序列的系统进化树,枝上数字为bootstrap 百分比。ST为来自山丘型流行区的样本,ZY和DG为来自两处湖沼型流行区的样本。
3 讨 论
日本血吸虫病流行区类型主要分为湖沼型流行区、洲滩型和山丘型。本研究选择了来自安徽省三个县(区)、两种流行区的日本血吸虫样本,对两个线粒体基因片段进行序列分析,从而初步探讨不同地区日本血吸虫群体的线粒体基因差异。从不同地区的纵向比较来看,无论是ND1基因还是COⅠ基因,湖沼型流行区样本的变异位点数、核苷酸多样性(Pi)、核苷酸差异平均数明显高于山丘型流行区。这可能是因为山丘型流行区环境相对封闭和保守,其间的生物群体发生基因交流的机会较少,发生的频率也更低,因此种群相对稳定;而湖沼型、水网型地区,水域面积广阔,加之不时爆发的夏季洪水,这些流行区的钉螺随水流迁移更频繁,导致日本血吸虫基因交流也更频繁。
日本血吸虫在中国分布的种系研究显示,不同地域的日本血吸虫在基因、表型、流行特征等方面不完全相同,学者认为可将其分为不同的地域株[14-16]。邱持平[14]研究发现湖南、安徽、湖北、四川及江苏的日本血吸虫株ND1基因和COⅠ基因存在两种基因型。郭凯文[15]依据线粒体COⅠ 和ND1基因构建的系统进化树显示中国大陆日本血吸虫分为三类:中国山区型地域株、湖沼江滩型地域株、湖北地域株。安徽省日本血吸虫病流行区以湖沼型为主,兼有水网型和山丘型。池州市石台县是安徽省内典型的山丘型血吸虫病流行区。石台地区的日本血吸虫具有和其他地区日本血吸虫不同的生物学特性,其主要保虫宿主是野鼠[17-18],且当地感染性钉螺具有夜间逸蚴的习性[19-20]。本研究首次依据线粒体COⅠ和ND1基因对石台地区日本血吸虫群体进行遗传进化研究,结果表明石台县日本血吸虫群体的COⅠ基因与枞阳县、大观区的样本之间具有一定的差异,而ND1基因没有发生明显的变化。依据本研究结果,尽管石台山区的日本血吸虫出现了流行病学和生物学上的差异,但是基因水平差异还尚不明显。这可能是因为三地均位于安徽省内,且三者之间地理位置较近。
本研究结果初步表明安徽省石台县山丘型日本血吸虫病流行区的日本血吸虫在线粒体基因水平与湖沼型流行区(安庆市大观区、铜陵市枞阳县)的日本血吸虫具有一定差异。基于石台县日本血吸虫保虫宿主和夜间逸蚴习性的特殊性,其基因差异是否会影响其对小鼠或者家兔的感染性和致病性等还有待进一步研究。
猜你喜欢
天津市教科院学报(2021年5期)2021-11-10
生物学通报(2021年9期)2021-07-01
华人时刊(2021年21期)2021-03-09
热带病与寄生虫学(2021年2期)2021-02-28
中国感染与化疗杂志(2020年1期)2020-01-12
家庭用药(2018年3期)2018-03-23
现代装饰(2017年9期)2017-05-25
中国医学创新(2017年7期)2017-03-31
江苏农业科学(2016年8期)2017-02-15
中国三峡(2016年6期)2017-01-15
