
河北省森林草原区草本植物叶功能性状种间和种内变异1)
2021-12-03 19:17冯家宝
东北林业大学学报 2021年1期
冯家宝
(河北省林木种质资源与森林保护重点实验室(河北农业大学), 保定,071000)
范顺祥 侯煜飞 剧永望 马天蕊 高英月 张志东
(中国农业大学) (河北省林木种质资源与森林保护重点实验室(河北农业大学))
植物功能性状是决定植物生长、定植、繁殖和存活的重要生理生态指标[1]。它不仅影响着植物个体的外在表现,同时也影响着植物群落的构成和生态功能。Diazd et al.[2]认为植物功能性状是联系物种、生境条件和生态功能的纽带。在种内和群落水平上了解功能性状之间的关系或权衡不仅有助于揭示物种对环境的适应策略和群落构建过程[3-5],而且能够更好的阐明生物多样性的维持机制[6]。
功能性状变异包括种内和种间变异。一般来讲,功能性状在种间存在很大的差异。因而,种间变异常被用于定义植物生长、功能策略等方面[7]。也有学者认为,种间变异对于群落构建的作用更大[8]。种内性状变异是由种群间的遗传差异、对环境梯度的可塑响应和个体发育变化引起的[4],是同种不同个体适应不同环境能力的体现。长期以来,有关植物功能性状的研究多以种间变异为主,对种内性状的研究较少[8-9]。然而,越来越多的证据表明,定量种内性状变异程度对于理解功能性状在群落水平变异及生态过程中发挥的作用越来越大[10]。基于此,结合种内和种间性状变异进行理解群落构建和生态过程就显得很有必要。
植物叶片在大气和陆地间碳交换方面发挥了关键的作用[11],其功能性状能够很好的反映物种对环境的响应、生活史权衡及资源获取策略[12]。不同的光合途径类型影响叶生理功能经济谱[13]。如相对于C3植物,C4草本植物通过叶结构的特殊变化促进光合作用,进而提高了光合效率和水分利用效率[13]。阐明C3和C4植物叶功能性状变异有助于更好的理解二者的光合机理和共存机制。
河北省塞罕坝地区处于华北典型森林-草原生态交错带,生态系统复杂多样,集森林、草原、草甸、沼泽为一体,物种多样性丰富,其中以草本植物为主,是河北省少有的物种多样性富集中心,具有重要的保护价值。特殊的生境条件和较高的草本物种多样性为研究草本植物叶功能性状变异和物种共存机制提供了良好的条件。本研究在景观水平进行分层随机取样的基础上,基于光合作用途径对草本植物进行了功能群的划分,通过比较5种功能性状的种间和种内变异,试图阐明研究区C3和C4草本植物主要功能性状的差异和相关性,为森林-草原生态交错带物种多样性保护提供科学依据。
1 研究区概况
研究区位于河北省承德市塞罕坝国家级自然保护区(42°22′~42°31′N,116°53′~117°31′E)。本区地貌是高原—波状丘陵—漫滩—接坝山地的组合体。气候属于温带半湿润气候与温带半干旱气候的过渡区。年均气温-1.5 ℃,极端最高气温30 ℃,极端最低气温-42.8 ℃,≥5 ℃的年积温1 930.7 ℃;年均降水量452.6 mm,蒸发量1 230 mm;植被由森林向草原过渡,草本植物物种多样性丰富,占到了所有物种的80%以上,主要由委陵菜(Potentillachinensis)、冰草(Agropyroncrsiatutm)、地榆(Sanguisorbaofficinalis)、金莲花(Trolliuschinensis)、披针叶苔草(Carexlanceolata)等物种构成[14]。
2 材料与方法
2.1 样地设置和样品采集
于2014年7—9月份在研究区进行群落学调查和样品采集。在整个研究区,按照斑块大小设置不同数量的调查样地。在空间上不连续的38个天然植被斑块(12个草本、11个灌木、15个天然林)中共调查草本样地184个,样地大小为1 m×1 m。记录草本植物的物种名、多度和盖度。基于功能性状测定方法[15],在每个样地内,选取盖度大于90%的所有物种,每个物种采集10株个体,测定计算完好成熟叶片的鲜质量。叶绿素质量分数测定,使用野外手持式叶绿素荧光仪进行测量。比叶面积测定,首先使用叶面积仪(LI-COR 3100C Area Meter, LI-COR, USA)测定叶面积,用电子天平测量叶片鲜质量,然后将叶片样品置于烘箱60 ℃烘干72 h后,称质量测定叶片干质量,比叶面积为叶片单位干质量所拥有的叶面积大小。叶片干物质质量分数测定,叶片样品的干质量/叶片鲜质量,它代表除去水分等因素后一片新鲜叶片中所含干物质的比例。全氮质量分数测定,叶片单位干质量中含有的氮的全量,烘干后的叶片样品用半微量凯氏定氮仪测定。全磷质量分数测定,叶片单位干质量中含有磷的全量,烘干后的叶片样品用钼锑抗比色法测定[16]。
2.2 数据分析
选取物种多度较高的优势种23种(C3植物13种,C4植物10种)(表1)进行植物功能性状种间和种内变异分析。采取Komogorov-Smirnov非参数检验方法比较C3和C4植物在5个不同功能性状上的差异。种内和种间植物功能性状的变异程度用变异系数来描述,其公式为:变异系数=(标准偏差÷平均值)×100%。利用皮尔逊相关分析分别检验C3和C4植物在种间和种内水平上不同功能性状间的相关性。运用R软件(3.3)中的“pwr”和“nlme”程序包完成以上统计检验,显著水平设定为P<0.05。

表1 C3和C4植物优势种科和属统计
3 结果与分析
3.1 C3和C4植物功能性状差异
5个功能性状叶绿素质量分数(D=0.528,P<0.01,D是Komogorov-Smirnov非参数检验中描述两组数据之间差异的统计量),叶片干物质质量分数(D=0.267,P<0.05),比叶面积(D=0.128,P<0.05),叶氮质量分数(D=0.610,P<0.01)和叶磷质量分数(D=0.507,P<0.01)在C3植物和C4植物间均存在显著性差异(表2)。C3植物的叶绿素质量分数((5.99±4.61)mg·g-1)、比叶面积((163.84±101.23)cm2·g-1)、全氮质量分数((15.85±2.73)g·kg-1)、全磷质量分数((0.14±0.04)g·kg-1)显著高于C4植物叶绿素质量分数(1.81±1.46)mg·g-1)、比叶面积((157.42±94.86)cm2·g-1)、全氮质量分数((13.05±2.96)g·kg-1)和全磷质量分数((0.09±0.04)g·kg-1)。与此相反,C3植物的叶干物质质量分数((0.32±0.13)g·g-1)则显著低于C4植物的((0.39±0.12)g·g-1)。

表2 C3和C4植物种的5种功能性性状差异
注;表中数据为平均值±标准差。
3.2 C3和C4植物种间和种内变异系数
无论在种间还是种内水平,C3植物和C4植物的5个性状均存在不同程度的变异。由表3可以看出,种间各性状的变异程度明显大于种内。C3植物和C4植物叶片干物质质量分数,叶氮、磷质量分数在种内变异系数比较接近,且均在20%左右。C3植物和C4植物叶绿素质量分数和比叶面积种内变异系数差异较大。C3植物和C4植物叶片干物质质量分数在种间变异系数比较接近(分别为56.37%和51.38%),比叶面积,叶绿素、叶氮、叶磷质量分数在种间变异系数均有较大差异。特别是比叶面积,叶氮、磷质量分数的种间变异系数的差异均在30%左右(C4植物大于C3植物)。无论是C3植物还是C4植物种间的变异系数均大于种内,除C4植物叶绿素质量分数、C3植物比叶面积差异不明显外,其他性状在种内、种间变异系数差异均在10%以上。
3.3 种间和种内水平上植物功能性状的相关性
C4植物各功能性状种间水平的相关性显著程度大部分高于种内水平,其中在种间水平上,比叶面积与叶片干物质质量分数呈极显著负相关,而与叶氮质量分数呈显著正相关(P<0.01)(表4);叶干物质质量分数会随着叶氮质量分数、叶磷质量分数增加而显著减少;叶磷质量分数与比叶面积、叶氮质量分数的相关关系显著(P<0.05)。而在种内水平上,只有比叶面积表现出了与叶片干物质质量分数、叶氮质量分数显著的相关关系(P<0.05)。C3植物各功能性状种内种间水平的相关性显著程度差异不大。无论在种间、种内水平上比叶面积均表现出与叶片干物质质量分数显著相关(P<0.05)。

表3 不同光合型植物功能性状种内和种间变异系数
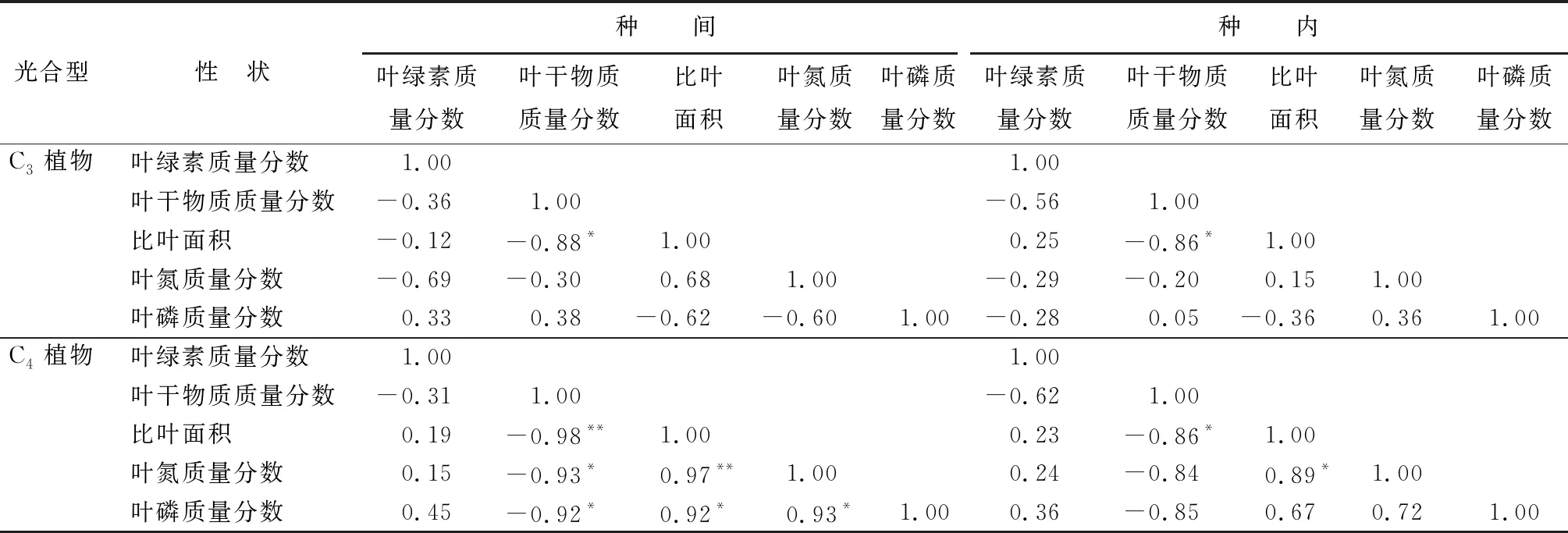
表4 种内、种间水平不同光合型植物功能性状相关性
4 结论与讨论
植物在进化与演替过程中经过一系列环境条件的筛选形成不同的功能性状组合和权衡来适应环境,进而来决定植物的生长策略[17]。本研究中C3植物和C4植物在5个植物功能性状(叶绿素质量分数、比叶面积、叶片干物质质量分数、叶氮质量分数、叶磷质量分数)上均表现出了显著的差异性,正是C3植物和C4植物为了适应环境,采用不同的适应机制及生存策略而产生的。
C4植物与C3植物相比,具有低呼吸、低CO2补偿点、高光饱和点、高的净光合速率和光量子效率等特点,同时对水分、氮素、光及CO2利用效率也较高[14]。本研究结果表明,C3植物的叶绿素质量分数,比叶面积,叶氮、磷质量分数均显著大于C4植物。叶绿素作为植物吸收光能的重要物质,在高等植物的光合作用中扮演着重要的角色,其质量分数的多少直接影响光合作用的强弱[18]。而叶氮、磷质量分数作为植物光合作用的协同元素,同样对光合作用进程有较大的影响。叶氮、磷质量分数,比叶面积,光合作用能力之间互相呈正相关[19],一般来说增加叶氮、磷质量分数能够显著提高叶片光合作用能力[20-21]。在相同的环境条件下,光合作用率较低的C3植物需要更多的叶绿素,叶氮、叶磷质量分数来维持光合作用以获取能量,这也侧面反映了植物通过增加或减少养分循环的方式来适应环境变化[22]。比叶面积的大小反映了植物对资源的获取、保存和释放能力,较大的比叶面积更有利于植物对光照的捕获和利用[23]。相对于C4植物,C3植物较大的比叶面积更容易接收光照来促进光合作用。与之相反,C3植物的叶干物质质量分数低于C4植物。可以解释为叶干物质质量分数作为植物对养分元素保有能力的体现[24],较大的叶干物质质量分数意味着植物叶片组织密度越大,其抵抗和耐受能力越强[25]。研究表明,C4植物适宜生长在高温干旱的环境下[26],植物为了增强抵抗能力,将更多的干物质投入到抵御不良环境中[27],因此,比C3植物有更高的干物质质量分数。此外,在不同研究中,植物叶片的叶干物质质量分数和比叶面积之间普遍表现出的负相关关系是两种生物量投资策略权衡的结果[28-29],本研究进一步证明了这个观点。
C3植物和C4植物生理生态上的差异导致了研究区内两类植物的功能性状在面对相同环境条件下表现出的差异性,从而形成了研究区多样性的分布格局。
植物功能性状的种间变异反映的是在不同生境下所产生的不同物种组成变化方式,其通常是受环境条件和遗传的差异影响而形成的[30]。许多研究表明,植物功能性状的分异主要表现在种间水平[19,31],本研究中,C3植物和C4植物的种间相关性和变异程度明显大于种内变异也体现了这一点。种内水平功能性状代表了物种个体基因造成的差异性及环境条件产生的形态差异[32]。本研究5种植物功能性状的种内变异系数为15%~40%,甚至某些性状变异程度已经接近种间变异。以往研究表明,如果种内变异较小就采取以平均值的形式代表物种的性状特征[33],这样的方式忽略了个体基因变异和环境异质性等产生的形态差异的影响。而李远智[34]在研究亚高寒草甸性状与群落构建机制时就发现,尽管种内变异较种间变异要小,但是种内性状变异对群落水平性状的梯度贡献却更高。这表明种内变异对群落构建有重要作用,在进行研究时同样不可忽视。只有充分考虑物种个体水平的功能性状差异才能更好的阐明物种共存和群落的构建机制[35],这一观点也被许多研究证实[30,36]。
在群落构建过程中,植物功能性状的筛选一般经历两个过程,生境过滤筛选出一类适合环境条件的相似的功能性状,而相似性限制又致使物种为了避免竞争和生态位重叠,发生性状趋异[37]。种内性状变异中比叶面积、叶绿素质量分数变异更大,是因为二者代表了植物个体在竞争资源中对个体产出策略以及对光合作用投资的变化,进而产生的一种减少竞争的行为。变异系数的大小可以表示性状的可塑性,较大变异系数表明,性状的可塑性较强,反之则表示较小的性状可塑性[38]。本研究中C4植物性状种间变异总体较大,表明C4植物更能适应变化的环境,具有较强的耐性和抗性,可能会成为演替后期的优势种。
本研究以塞罕坝自然保护区中主要草本植被为研究对象,在划分功能群的基础上,进行了5个主要功能性状的种间和种内变异分析。5个植物功能性状都表现出了显著差异性,C3植物的叶绿素质量分数、比叶面积、叶氮质量分数、叶磷质量分数均大于C4植物。种间变异系数较大的C4植物对环境的耐受性更高,可以成为恢复退化草地的重要手段。从总体来看,种间变异系数较大,表明种间变异仍是植物功能性状分异的主要驱动因素,但是种内变异同样对群落构建有重要作用,依旧不可忽视。本研究结果有助于理解森林草原过渡区草本植物功能性状之间的关系或权衡,揭示物种对环境的适应策略和群落构建过程,可为研究区草本植物物种多样性保护提供理论依据。
猜你喜欢
河南师范大学学报(自然科学版)(2022年5期)2022-08-08
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02
趣味(数学)(2020年4期)2020-07-27
支部建设(2020年15期)2020-07-08
新疆农业科学(2020年1期)2020-02-14
蔬菜(2018年5期)2018-05-17
杂草学报(2015年2期)2016-01-04
中国麻业科学(2015年5期)2015-12-28
百科知识(2015年18期)2015-09-10
中国海洋大学学报(自然科学版)(2014年12期)2014-02-28
