
黄淮麦区(南片)小麦新品系脂肪氧化酶活性分析及其等位基因检测
2021-12-08 03:32张星宇张福彦宋佳静宋全昊赵立尚朱统泉梁宝萍宋晓朋
麦类作物学报 2021年10期
陈 杰,张星宇,张福彦,白 冬,宋佳静,宋全昊,金 艳,赵立尚,朱统泉,王 勇,梁宝萍,李 艳,宋晓朋
(1.河南省驻马店市农业科学院,河南驻马店 463000; 2.河南农业大学农学院,河南郑州 450002;3.河南省科学院同位素研究所有限责任公司/河南省核农学重点实验室,河南郑州 450015)
面粉色泽是评价小麦加工品质的重要指标之一,随着过氧化苯甲酰等面粉增白剂的禁用,选育自然白的小麦品种成为当务之急。面粉色泽不仅受到类胡萝卜素、黄色素等色素类物质的影响,同时还受到多酚氧化酶、过氧化物酶等酶类物质的影响。Loiseau等[1]和Dong等[2]研究表明,小麦籽粒中脂肪氧化酶(lipoxygenase,Lox)可氧化降解类胡萝卜素进而使面粉增白,因此,研究小麦籽粒中的Lox对改良小麦面粉色泽具有重要意义。
环境和基因型都能够影响Lox的活性,但基因型对Lox活性的影响较大[3-4]。小麦籽粒Lox活性是受多基因调控的数量性状。前人通过定位发现,小麦1A、4B、5D和7B 染色体上均存在影响Lox活性的基因位点,并发现了与其紧密连锁的SSR标记(Xwmc312、Xbcd1262、Xgwm251和Xksud2a)[5-7]。研究表明,小麦4B染色体上的基因位点对小麦籽粒Lox活性的影响较大[5-9]。近年来,随着小麦生物信息学的发展和同源克隆技术的应用,普通小麦Lox活性基因的克隆以及功能标记的开发也取得了快速的发展。Geng等[8]利用同源克隆的方法克隆了4B染色体上的TaLox-B1基因, 并根据等位变异序列之间的差异,开发设计了两对共显性互补功能标记Lox16和Lox18,用来检测TaLox-B1位点上的等位基因TaLox-B1a和TaLox-B1b。标记Lox16和Lox18已经在新疆、陕西、黑龙江和宁夏小麦材料检测中得到了应用[10-13]。随后,Zhang等[9]利用同源克隆的方法克隆了4B染色体上的TaLox-B2和TaLox-B3基因, 并根据等位变异序列之间的差异,开发设计了一对共显性功能标记Lox-B23,用来检测TaLox-B2和TaLox-B3位点上的等位基因TaLox-B2a、TaLox-B2b和TaLox-B3a、TaLox-B3b。标记Lox-B23已经在河南和新疆小麦材料检测中得到了应用[14-15]。
黄淮麦区(南片)小麦生产在保障我国粮食安全中发挥着重要作用,该区小麦产量、抗性、面筋品质等相关性状的遗传改良已经有了很大进步,但有关小麦籽粒Lox活性的改良尚显不足。本研究以94份黄淮麦区(南片)小麦新品系为材料,利用功能标记Lox16、Lox18和Lox-B23对其TaLox-B1、TaLox-B2和TaLox-B3位点上的等位基因进行检测,同时利用分光光度计对供试材料的Lox活性进行测定,分析TaLox-B1、TaLox-B2和TaLox-B3位点上等位基因的分布情况以及不同等位基因和Lox活性之间的关系,以期为小麦Lox活性分子标记的辅助育种提供参考。
1 材料与方法
1.1 供试材料
试验材料为2020-2021年度黄淮冬麦区南片水地组区域试验的小麦新品系,剔除组别间重复的对照品种,合计94份(表1)。其中来源于河南、安徽、陕西、江苏和北京的材料分别有52、16、12、10和4份。这些材料都是在黄淮麦区(南片)23个试验点经过两年品种比较试验晋级上来的材料,可以很客观地反映本麦区当前的小麦育种水平。

(续表1 Continued table 1)
1.2 脂肪氧化酶(Lox)活性的测定
利用UV-2601型分光光度计(北京瑞利分析仪器有限公司生产)测定供试材料的Lox活性,具体步骤参照吴培培等[4]的方法。
1.3 等位基因的分子检测
参照陈 杰等[16]的方法提取参试材料的基因组DNA。用Lox16和Lox18标记检测参试材料TaLox-B2位点的等位基因,扩增出489 bp 条带的材料记为TaLox-B1a类型,扩增出791 bp条带的材料记为TaLox-B1b类型;用 Lox-B23标记检测参试材料TaLox-B2和TaLox-B3位点的等位基因,扩增出788 bp 条带的材料记为TaLox-B2a类型,扩增出660 bp条带的材料记为TaLox-B2b类型,扩增出677 bp 条带的材料记为TaLox-B3a类型, 未扩增出任何带型的记为TaLox-B3b类型。所用标记的引物序列及退火温度等信息见表2。

表2 Lox不同位点等位基因检测引物信息Table 2 Primers for identification of different Lox alleles
PCR反应体系和扩增程序参照陈 杰等[16]的方法。标记Lox16和Lox18的PCR扩增产物用2%的琼脂糖凝胶电泳分离,标记Lox-B23的PCR扩增产物用3%的琼脂糖凝胶电泳分离,电泳缓冲体系为1×TBE Buffer溶液,电压设置为160 V,电泳时间为40 min,电泳结束后用0.5%的溴化乙锭(EB)染色,用凝胶成像系统 (Protein Simple,美国)查看PCR扩增带型,并拍照保存。
1.4 数据分析
用Microsoft Excel 2003进行数据分析,用SPSS 18.0进行方差分析。
2 结果与分析
2.1 供试材料籽粒Lox活性的分布
对94份供试材料的Lox活性进行测定,结果(表1)表明,供试材料Lox活性的总体平均值为71.47 AU·min-1·g-1,标准差为5.49,变幅为59.8~83.1 AU·min-1·g-1。Lox活性低于70 AU·min-1·g-1的材料有38 份,占比40.4%;Lox活性在70~75 AU·min-1·g-1的材料有29份,占比30.9%;Lox活性值高于75 AU·min-1·g-1的材料有27份,占比 28.7%。其中,有6份材料(冠麦12、漯麦66、淮麦40、泉麦39、周麦36号和周麦49号)的Lox活性高于80 AU·min-1·g-1。有12份材料(西农172、涡麦179、许科10号、郑麦20、徐麦15019、新农9799、科麦1609、安科1704、丰工40、丰工38、淮麦701和平安918)的Lox活性低于65 AU·min-1·g-1。说明黄淮麦区(南片)大部分新育成小麦材料均以中等Lox活性为主。
2.2 TaLox-B1、TaLox-B2和TaLox-B3位点等位基因的分布及其与Lox活性的关系
用标记Lox-16和Lox-18对供试材料的TaLox-B1位点进行等位基因检测,部分材料的检测结果如图1所示。94份供试材料中,淮核16174、瑞华麦519、漯麦68等17份材料可以扩增出489 bp的条带,说明这些材料含有TaLox-B1a等位基因,分布频率为18.1%;淮核16132、华麦15080、保丰1707等77份材料可以扩增出791 bp的条带,说明这些材料含有TaLox-B1b等位基因,分布频率为 81.9%。从以上检测结果分析可知,94份供试材料的TaLox-B1位点以TaLox-B1b等位基因类型分布为主。
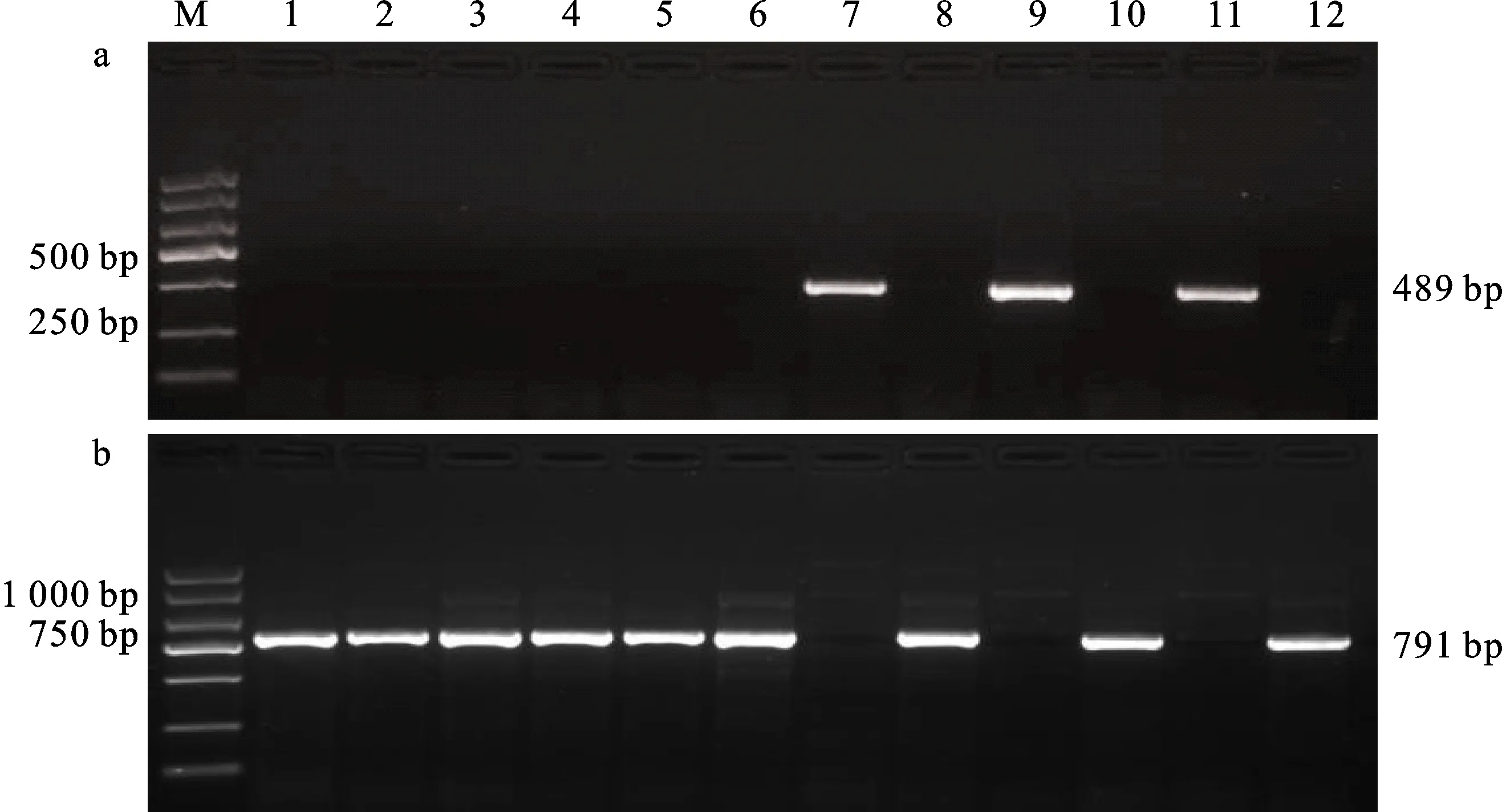
a:Lox-16标记的扩增结果;b:Lox-18标记的扩增结果;M:Marker;1:淮核16132;2:华麦15080;3:保丰1707;4:淮麦701;5:瑞华556;6:保麦1633;7:淮核16174;8:徐麦15019;9:瑞华麦519;10:淮麦40;11:漯麦68;12:驻麦586。图2同。a:Amplification result by Lox-16; b:Amplification result by Lox-18; M:Marker; 1:Huaihe 16132; 2:Huamai 15080:3:Baofeng 1707; 4:Huaimai 701; 5:Ruihua 556; 6:Baomai 1633; 7:Huaihe 16174; 8:Xumai 15019; 9:Ruihuamai 519; 10:Huaimai 40; 11:Luomai 68; 12:Zhumai 586. The same in figure 2.图1 Lox-16和Lox-18标记对部分供试小麦品种TaLox-B1位点的扩增结果Fig.1 Amplification results of TaLox-B1 by markers Lox 16 and Lox 18 in some wheat cultivars
用标记Lox-B23对供试材料的TaLox-B2和TaLox-B3位点进行等位基因检测,部分材料的检测结果如图2所示。在TaLox-B2位点,淮核16132、华麦15080、保丰1707等83份材料可以扩增出788 bp的条带,说明这些材料含有TaLox-B2a等位基因,分布频率为88.3%;徐麦15019、涡麦179、许科10号等11份材料可以扩增出660 bp的条带,说明这些材料含有TaLox-B2b等位基因,分布频率为11.7%;在TaLox-B3位点,华麦15080、保丰1707、瑞华556等63份材料可以扩增出677 bp的条带,说明这些材料含有TaLox-B3a等位基因,分布频率为67.0%;淮核16132、淮麦701、淮核16174等31份材料未扩增出任何条带,说明这些材料含有TaLox-B3b等位基因,分布频率为33.0%。从以上检测结果分析可知,94份供试材料的TaLox-B2和TaLox-B3位点分别以TaLox-B2a和TaLox-B3a等位基因类型分布为主。
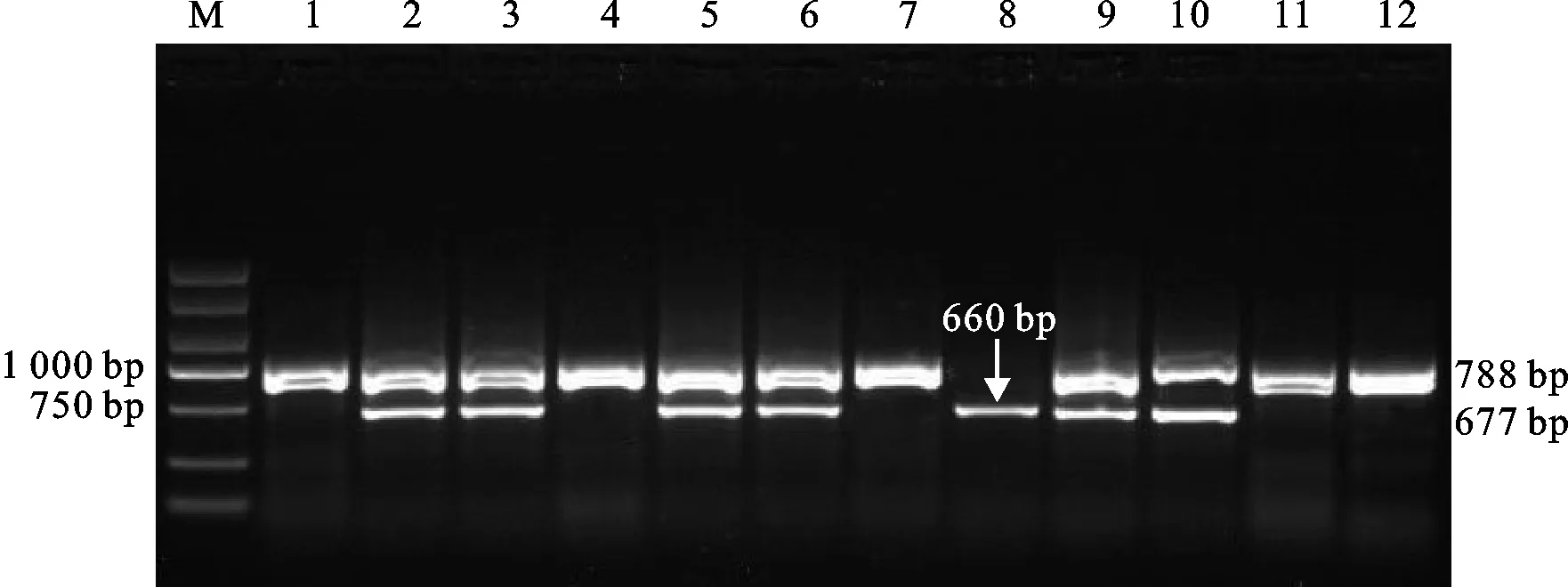
图2 Lox-B23标记对部分供试小麦品种TaLox-B2和TaLox-B3位点的扩增结果Fig.2 Amplification results of TaLox-B2 and TaLox-B3 by marker Lox-B23 in some wheat cultivars
从表3可以看出,在TaLox-B1、TaLox-B2和TaLox-B3位点,分别检测到两种等位基因,且两种等位基因之间的Lox活性差异均达到显著水平,TaLox-B1a、TaLox-B2a和TaLox-B3a等位基因与高Lox活性相关,TaLox-B1b、TaLox-B2b和TaLox-B3b等位基因与低Lox活性相关。
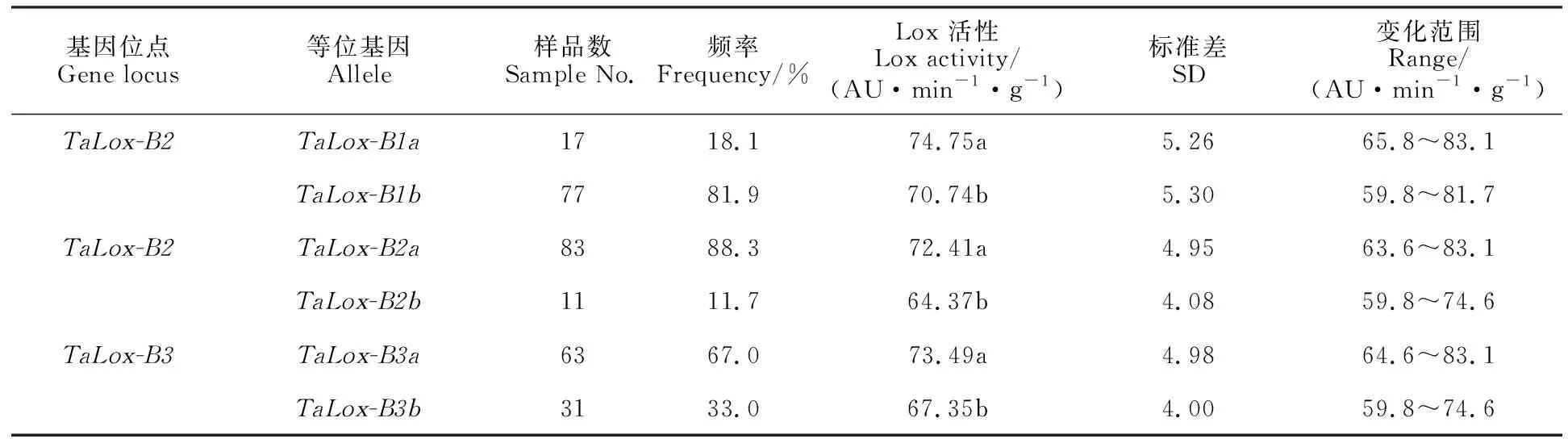
表3 不同位点等位基因对Lox活性的影响Table 3 The effect of different alleles on Lox activity
2.3 不同等位基因组合的分布及其与Lox活性的关系
94份供试材料中,TaLox-B1、TaLox-B2和TaLox-B3位点共检测到六种类型的等位基因组合(表4)。其中含有TaLox-B1b/TaLox-B2a/TaLox-B3a等位基因组合的材料最多,有53份,分布频率为56.4%;含有TaLox-B1a/TaLox-B2b/TaLox-B3b等位基因组合的材料最少,仅有2份,分布频率为2.1%;其余四种类型等位基因组合的分布频率由高到低依次为TaLox-B1b/TaLox-B2a/TaLox-B3b、TaLox-B1a/TaLox-B2a/TaLox-B3a、TaLox-B1b/TaLox-B2b/TaLox-B3b和TaLox-B1a/TaLox-B2a/TaLox-B3b,分布频率分别为16.0%、10.6%、9.6%和 5.3%。从以上检测结果分析可知,94份供试材料以TaLox-B1b/TaLox-B2a/TaLox-B3a等位基因组合类型分布为主。
进一步分析不同等位基因组合对小麦籽粒Lox活性的影响,结果(表4)可以看出,含有TaLox-B1a/TaLox-B2a/TaLox-B3a等位基因组合类型材料的Lox活性最高,与其余五种类型差异显著。含有TaLox-B1b/TaLox-B2b/TaLox-B3b等位基因组合类型材料的Lox活性最低,与其余五种类型也差异显著。其余四种等位基因组合类型中,含有TaLox-B1b/TaLox-B2a/TaLox-B3b等位基因组合类型材料的Lox活性最低,与其余三种类型差异显著。这表明TaLox-B1a/TaLox-B2a/TaLox-B3a等位基因组合与高Lox活性相关,TaLox-B1b/TaLox-B2b/TaLox-B3b等位基因组合与低Lox活性相关,TaLox-B1b/TaLox-B2a/TaLox-B3b等位基因组合与中低Lox活性相关,其余三种等位基因组合与中等Lox活性相关。

表4 不同等位基因组合对Lox活性的影响Table 4 Effect of different allele combinations on Lox activity
2.4 Lox活性基因等位变异在不同省份(直辖市)供试材料中的分布频率
从表5可知,与高Lox活性相关的TaLox-B1a等位基因在来源于安徽、河南、江苏、陕西和北京小麦新品系中的分布频率分别为18.7%、 17.3%、20.0%、16.7%和25.0%,均低于与低Lox活性相关的TaLox-B1b等位基因;与高Lox活性相关的TaLox-B2a等位基因在来源于安徽、河南、江苏、陕西和北京小麦新品系中的分布频率分别为75.0%、 92.3%、90.0%、91.7%和 75.0%,均高于与低Lox活性相关的TaLox-B2b等位基因;与高Lox活性相关的TaLox-B3a等位基因在来源于安徽、河南、江苏、陕西和北京小麦新品系中的分布频率分别为 62.5%、67.3%、 60.0%、75.0%和75.0%,均高于与低Lox活性相关的TaLox-B3b等位基因。说明来源于安徽、河南、江苏、陕西和北京小麦新品种均以TaLox-B1b、TaLox-B2a和TaLox-B3a分布为主。

表5 LOX活性等位基因在不同省份材料中的分布频率Table 5 Frequency of LOX allelesin different provinces
与高Lox活性相关的TaLox-B1a/TaLox-B2a/TaLox-B3a等位基因组合在来源于安徽、河南、江苏、陕西和北京小麦新品系中的分布频率分别为6.3%、11.5%、10.0%、8.3%和25.0%;与低Lox活性相关的TaLox-B1b/TaLox-B2b/TaLox-B3b等位基因组合在来源于安徽、河南、江苏、陕西和北京小麦新品系中的分布频率分别为18.8%、5.8%、10.0%、8.3%和25.0%。说明来源于安徽、河南、江苏、陕西和北京小麦新品系均以中等Lox活性相关的等位基因组合分布为主。
3 讨 论
了解Lox活性基因位点不同等位基因的分布情况,可为Lox活性的分子遗传改良提供参考。前人研究表明,TaLox-B1b等位基因在新疆、陕西、黑龙江和宁夏小麦中的分布频率分别为75.90%、77.46%、88.98%和84.44%[10-13]。本研究结果表明,TaLox-B1b等位基因在黄淮麦区(南片)及其组成省份安徽、河南、江苏、陕西小麦新品系中的分布频率分别为81.9%、 81.3%、 82.7%、80.0%和83.3%。出现这种现象的原因可能是TaLox-B1b等位基因与低Lox活性相关,而低Lox活性的小麦较耐储藏[17],而高Lox活性可以使面粉增白,各地育种家在耐储藏和面粉增白两个性状上有目的地选择了前者。张福彦等[14]利用标记Lox-B23对122份河南小麦TaLox-B2和TaLox-B3位点进行等位基因检测,结果表明,TaLox-B2a和TaLox-B3a等位基因的分布频率分别为57.4%和41.8%。白 璐等[15]利用标记Lox-B23对123份新疆小麦TaLox-B2和TaLox-B3位点进行等位基因检测,结果表明,TaLox-B2a和TaLox-B3a等位基因的分布频率分别为26.8%和77.2%。本研究检测结果表明,TaLox-B2a和TaLox-B3a等位基因在94份黄淮麦区(南片)小麦新品系的分布频率分别为 88.3%和67.0%。综合以上数据可知,TaLox-B2a和TaLox-B3a等位基因在新疆麦区和黄淮南片麦区的分布频率存在很大的差异,这可能是材料来源不同所致。
本研究通过对94份供试小麦材料Lox活性和基因型关联分析可知,含有TaLox-B1a、TaLox-B2a和TaLox-B3a等位基因材料的Lox活性均显著高于其对应位点上含有TaLox-B1b、TaLox-B2b和TaLox-B3b等位基因的材料。这与Geng等[8]和Zhang等[9]的研究结果一致,验证了TaLox-B1a、TaLox-B2a和TaLox-B3a等位基因与高Lox活性相关,TaLox-B1b、TaLox-B2b和TaLox-B3b等位基因与低Lox活性相关,这也验证了标记Lox16、Lox18和Lox-B23可以作为Lox活性分子标记辅助育种的有效工具。但本研究也发现,部分携带与高Lox活性相关的TaLox-B1a/TaLox-B2a/TaLox-B3a等位基因组合小麦材料的表型值低于其他等位基因组合类型,造成这种情况的原因可能有两个,一是生态环境的影响,王 慧等[3]和吴培培等[4]研究表明,不同生态环境对小麦Lox活性存在影响,但生态环境的影响效应低于基因型的影响效应;二是小麦其他染色体上也存在影响籽粒Lox活性的基因位点,Feng等[6]和Garbus等[7]研究表明,普通小麦1A、5D和7B染色体也存在影响Lox活性的基因位点。另外,本研究中冠麦12、漯麦66和周麦49号不仅Lox活性高于80 AU·min-1·g-1,而且均携带与高Lox活性相关的TaLox-B1a/TaLox-B2a/TaLox-B3a等位基因组合,可作为选育高Lox活性和亮白面粉品种的亲本资源加以应用,Lox16、Lox18和Lox-B23三个标记可作为小麦Lox品种改良辅助选择的有效工具进一步利用。
猜你喜欢
分子催化(2022年1期)2022-11-02
河南农业·综合版(2022年2期)2022-03-18
河南农业(2022年2期)2022-03-14
河南农业·综合版(2021年7期)2021-08-23
智慧健康(2021年17期)2021-07-30
河南农业(2021年7期)2021-07-30
烟草科技(2021年6期)2021-06-24
生物学教学(2018年4期)2018-11-29
新课程·下旬(2018年9期)2018-11-14
电脑知识与技术(2018年19期)2018-11-01
