
平潭岛海岸自然植被物种和功能多样性特征
2021-12-11 04:36管诗敏郑心炫吴雅华李岩林杨桂梅杨钰华王伟耀黄柳菁
西南林业大学学报 2021年6期
管诗敏 郑心炫 吴雅华 李岩林 杨桂梅 杨钰华 王伟耀 黄柳菁
(福建农林大学园林学院,福建 福州 350002)
海岸植被作为海岸生态系统的有机部分,能够在海岸侵蚀、赤潮、海风、盐雾、高温等恶劣条件下生存[1],对抵抗自然灾害、维护海岸基础、分解污染物等方面有极大的生态价值[2]。然而,由于自然灾害加剧、旅游开发、养殖业发展、人工防护林的覆盖,海岸生态环境遭到极大地破坏,海岸自然植被种类、数量及分布范围大幅度下降[3],造成生物多样性下降。因此,急需对海岸自然植被进行生物多样性研究以促进海岸生态系统的保护及海岸植被恢复。
有学者认为生物多样性保护研究的关键是物种多样性[4],但有学者认为功能多样性能更有效的实现生物多样性保护[5]。物种共存机制的关键或与物种多样性及功能多样性有关,评估和管理生态系统的重点之一也与这两者有关[6]。目前物种多样性及功能多样性研究的区域主要集中在森林生态系统[7-9]、草地生态系统[10],少量海岛群落研究[11-12],海岸生态系统尤其是海岛的海岸植被研究鲜见报道。海岛因为其特殊的地理位置,同时受到大陆性气候和海洋性气候的影响,形成了相对独立且完整的生态系统[13],海岛面积有限,因此海岸作为海岛内陆与海洋交接的过渡带,其屏障作用尤为重要。
基于以上背景,本研究以平潭岛海岸原生植被中4种常见植被类型为研究对象,选取5个指数代表物种多样性、3个指数代表功能多样性,分析4种植被类型的物种多样性和功能多样性特征及其与环境因子的关系,进而丰富海岸植被的生物多样性保护理论研究,为海岸生态系统的保护管理提供参考。
1 研究区概况
平潭岛地处中国东南部沿海地区(25°15′~25°45′N,119°32′~120°01′E),位于福建省综合试验区内,是福建省第一大岛,陆地面积371.91 km2,海域面积6 000多km2,海岸线长399.82 km。该岛属南亚热带海洋季风性气候,夏热冬暖,年均气温19.6 ℃,最热月7—8月,月平均气温27.0~28.2 ℃;最冷月2月,月平均气温9.7~10.8 ℃。旱雨季节明显,雨季3—6月,年均降水量(1 172 mm)小于年均蒸发量(1 300 mm),属福建省少雨区。地势南北高、中部低,最高海拔为434.6 m。海岸线蜿蜒绵亘,海岸类型主要为基岩侵蚀海岸、红土侵蚀海岸、沙质塘积海岸等。植被多为逆行演替的人工植被,种类少,群落结构单纯。森林植被有台湾相思(Acacia confusa)、黑松(Pinus thunbergii)、木麻黄(Casu-arina equisetifolia)等植被,滨海植被有滨柃(Eurya emarginata)、海边月见草(Oenothera drummondii)、海滨藜(Atriplex maximowicziana)等植被。
2 研究方法
2.1 调查方法
2019年4—7月通过踏查,参考吴沙沙等[14]、郑俊鸣等[15]、《海南省七洲列岛的植物与植被》[16]对海岛植被的分类,沿海岸线选取不同自然植被类型,进行群落调查,共设置64个5 m×5 m样方,包括:海岸丘陵灌丛20个、海岸基岩灌丛12个、海岸基岩草丛8个、砂生草甸24个。记录调查样方内所有灌木和草本的物种名称、数量、盖度、生长型、高度等特征。测定环境因子,包括海拔、坡度、坡向、土壤容重、土壤水分、土壤含盐率、pH值。
2.2 测定方法
每个样方使用梅花五点法取5个直径为5 cm,深度为20 cm的土样。同时在刚取过土的5个样点取土混合均匀,再用另一个带有编号的自封袋装入500 g左右的土壤,送回实验室内测定pH值。在样方中心位置利用环刀取土样,后装入带有编号的自封袋中,用于土壤容重和含水率的测定。通过GPS手持定位仪测定经纬度、海拔高度、坡向、坡位、坡度。
2.3 分析方法
2.3.1 物种多样性计算
本研究选用了5种常用的物种多样性指数:Patrick丰富度(R0) 、Shannon-Weiner多样性指数(H′) 、Pielou均匀度指数(J)、Simpson优势度指数(λ)、和Jaccard相似性系数(Cj),其计算公式如下:
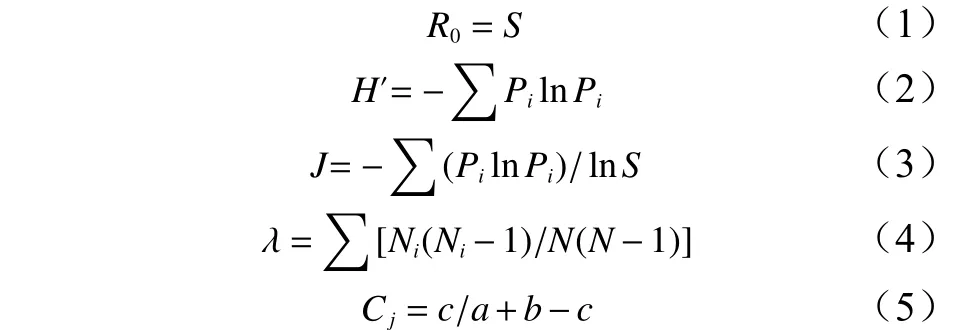
式中:S为样地中物种总数;N为所有物种总个体数;Ni是第i种的个体数;Pi为第i种的个体比例,i=1, 2, 3, ···,S。a、b分别为各自样方中的物种数;c为2个样方数中同时出现的物种数量。
2.3.2 功能多样性计算
功能多样性分为3个部分:功能丰富度、功能均匀度和功能离散度。功能丰富度选用功能性状平均距离指数(MFAD);功能均匀度选用多维功能均匀度指数(FEve)来表示;功能离散度选用常见的二次熵指数(Rao)来表示。计算公式如下:
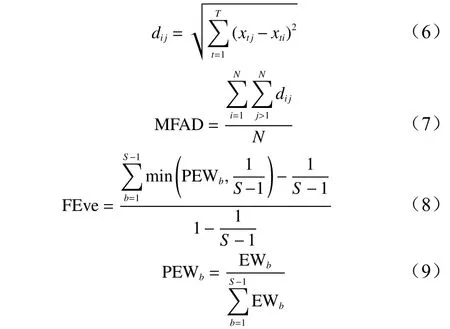
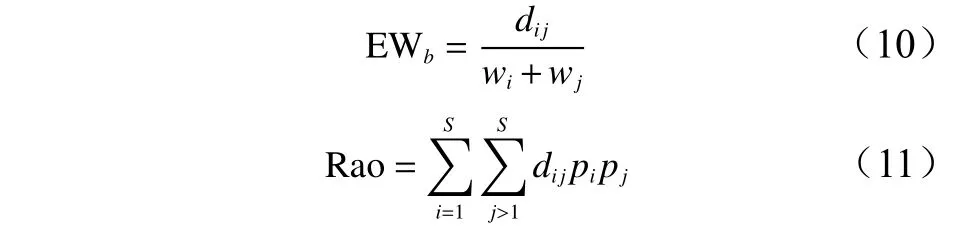
式中:T为性状数量,Ni物种i的个体数量,N为样方中所有物种的个体总数,xti和xtj分别为物种i和物种j性状t值,dij为物种i与物种j间的欧几里得度量。S为物种数目,PEWb为局部加权平均均匀度,EWb为加权平均的均匀度,wi和wj为物种i和物种j的相对多度。wi和wj分别为物种i和物种j的相对多度。pi和pj分别为物种i和物种j的个体数占群落总物种个体数的比例。
将植物功能性状中的名称数据进行赋值(表1),然后利用R语言3.6.2程序FD软件包中的dbFD函数计算以上3个功能多样性指数。

表1 植物功能性状特征类型Table 1 Types of plant functional traits
2.3.3 数据分析
通过Excel 2010进行数据初步整理,计算以上指数,经正态分布检验后,利用SPSS 25.0统计分析软件对以上2类指数进行单因素方差分析、Person相关性分析。选用 CANOCO 5.0 软件建立物种多样性−环境、功能多样性−环境矩阵进行去趋势对应分析(DCA),排序轴梯度长度为0.4、0.7,数据适用于线性模型。故采用冗余分析方法(RDA)分析环境因子对物种多样性和功能多样性的影响。
3 结果与分析
3.1 物种组成及相似性比较
海岸不同植被类型植物统计结果见表2。由表2可知,在4种植被类型中共记录107种植物,隶属于59科,103属;生长型以草本植物为主,共67种,占比62.6%,灌木共40种,占比37.3%。其中海岸丘陵灌丛物种数最高,共记录87个物种,隶属于49科、74属,分别占总记录的81.3%、83.%、71.8%;海岸基岩草丛次之,共记录31个物种,隶属于18科、29属,分别占总记录的29.0%、30.5%、28.2%;海岸基岩灌丛共记录26个物种,隶属于15科、23属,分别占总记录24.3%、25.4%、23.3%;砂生草甸物种数最少,共记录21个物种,隶属于15科、21属,分别占总记录19.6%、25.4%、20.4%。

表2 海岸不同植被类型植物统计表Table 2 Statistics of family, genus and species of different vegetation types on the coast
通过对平潭岛海岸4个不同植被类型的物种相似性分析,结果表明(表3):大部分植被类型间Jaccard物种相似性系数在0.00~0.25间,属于极不相似水平,其中丘陵灌丛与砂生草甸之间的Jaccard物种相似性系数为0.07,相似度最低;仅有基岩灌丛与基岩草丛的Jaccard物种相似性最高,为0.54,属于中等相似水平。
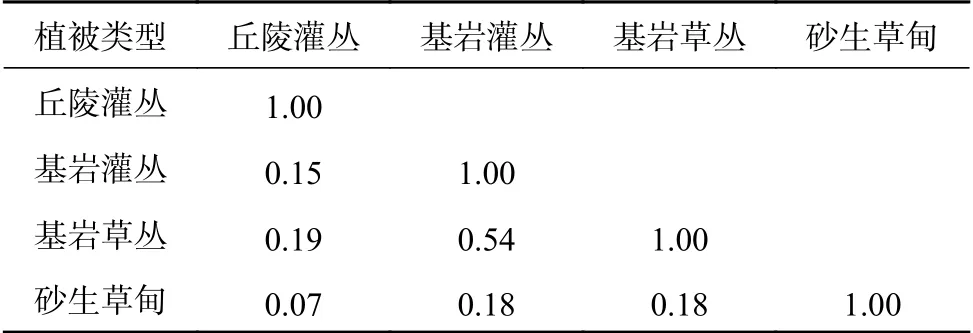
表3 海岸不同植被类型间相似性比较Table 3 Similarity comparison among different vegetation types on the coast
3.2 不同海岸植被类型的物种多样性
通过对平潭岛海岸4个不同植被类型的物种多样性指数分析,结果表明(图1):丘陵灌丛的丰富度、Shannon-Wiener多样性指数、Pielou均匀度指数高于其他3类且存在显著性差异(P<0.05)。4类植被类型彼此间的丰富度存在显著性差异(P<0.05);Shannon-Wiener多样性指数中基岩灌丛、基岩草丛与砂生草甸存在显著差异,基岩灌丛和基岩草丛没有显著性差异;均匀度指数中基岩灌丛、基岩草丛、砂生草甸没有显著性差异;Simpson优势度指数中砂生草甸均与其他3类且存在显著性差异(P<0.05),丘陵灌丛与基岩灌丛存在显著性差异,基岩灌丛与基岩草丛没有显著性差异。
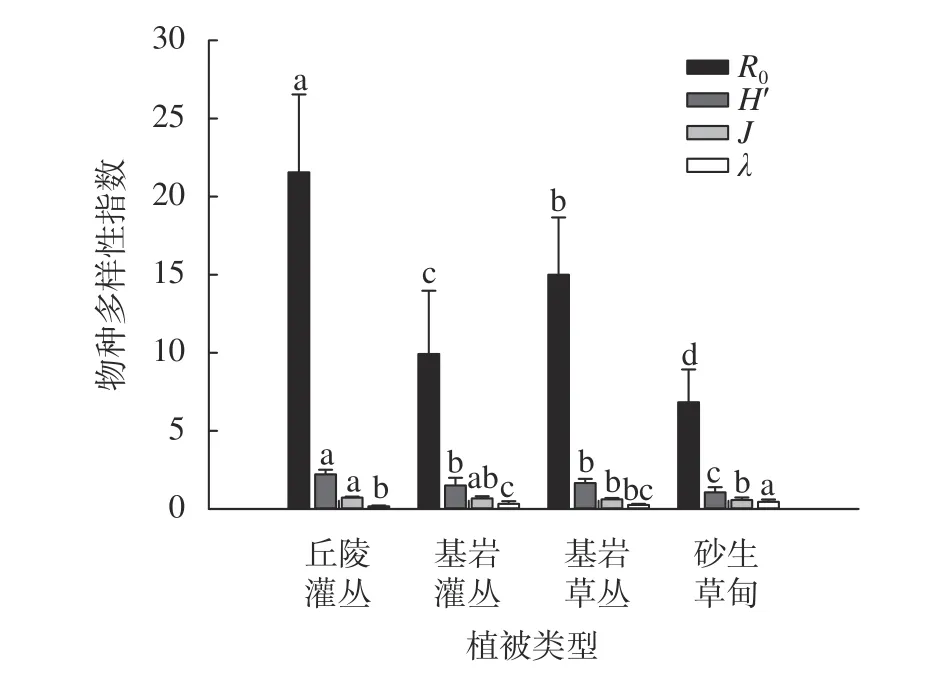
图1 海岸不同植被类型物种多样性Fig. 1 Species diversity of different vegetation types on the coast
3.3 不同植被类型的功能多样性
通过对平潭岛海岸4个不同植被类型的功能多样性分析,结果表明(图2):丘陵灌丛的功能丰富度指数最高,且丘陵灌丛的功能性状平均距离(MFAD)与基岩灌丛、基岩草丛、砂生草甸存在显著性差异(P<0.05),基岩灌丛、基岩草丛、砂生草甸之间没有显著性差异;丘陵灌丛的功能均匀度最高,且丘陵灌丛、基岩灌丛的多维功能均匀度指数(FEve)与基岩草丛、砂生草甸存在显著性差异(P<0.05);基岩灌丛的功能离散度最高,且丘陵灌丛、基岩灌丛的二次熵指数(Rao)与砂生草甸存在显著性差异(P<0.05),与基岩草丛没有达到显著性差异。
3.4 功能多样性和物种多样性间的相关性分析
对不同植被类型的所有样地物种多样性指数和功能多样性指数进行相关性分析(表4),代表功能多样性的3个指数间呈现极显著正相关关系(P<0.01),代表物种多样性的4个指数间也呈现极显著相关关系(P<0.01);而物种多样性和功能多样性的关系中,功能性状平均距离指数(MFAD)与Patrick丰富度、Shannon-Wiener多样性指数呈极显著正相关(P<0.01),与Pielou均匀度指数、Simpson优势度指数无显著相关关系;功能均匀度选用多维功能均匀度指数(FEve)与Patrick丰富度、Shannon-Wiener多样性指数呈极显著正相关(P<0.01),与Pielou均匀度指数呈显著正相关(P<0.05),与Simpson优势度指数呈显著负相关(P<0.05);二次熵指数(Rao)与Shannon-Wiener多样性指数呈显著正相关(P<0.05),与Pielou均匀度指数呈极显著正相关(P<0.01),与Patrick丰富度无显著相关关系。
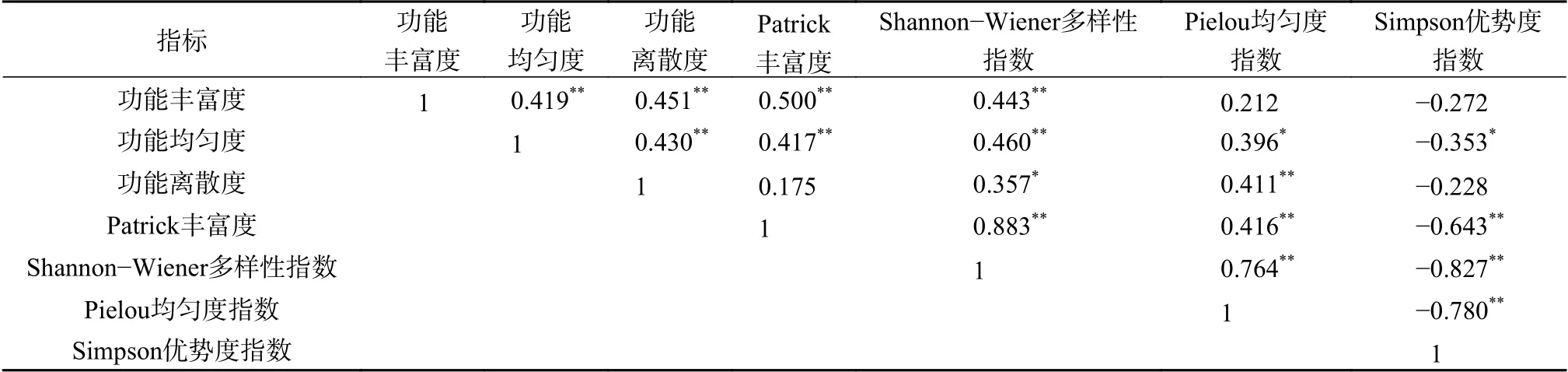
表4 物种多样性指数和功能多样性指数间Person相关性分析Table 4 Person correlation coefficients of functional diversity and species diversity
3.5 环境因子对物种多样性和功能多样性的影响
由图3可知,物种多样性指数与环境因子排序图中前2个主成分累计特征值占99.28%,调整后解释变量为21.8%(P<0.05);功能多样性指数与环境因子排序图中前2个主成分累计特征值占98.99%,调整后解释变量为30.5%(P<0.05)。影响物种多样性的环境因子主要是土壤含盐率、含水量、坡度,解释量分别为33.9%、32%、22%。物种丰富度和Shannon-Winner多样性指数与土壤容重、海拔呈正相关,与土壤含盐率、坡向、坡度呈负相关;Pielou均匀度指数随着土壤含盐率及含水量增加而降低;Simpson优势度指数随着土壤含盐率、含水量、坡度增加而增加。在功能多样性与环境因子的关系中,土壤容重、坡向、坡度、海拔是主要影响因素,解释量分别为31.9%、14.6%、18.2%、24.3%。MAFD、FEve与土壤含水量、土壤容重呈正相关,与坡向呈负相关;Rao与土壤含盐率及pH呈正相关,与坡度、海拔呈负相关。

图3 环境因子与物种多样性和功能多样性的关系Fig. 3 Relationship between environmental factors and species diversity or functional diversity
4 结论与讨论
4.1 讨论
4.1.1 不同植被类型的物种多样性
不同植被类型环境盐度和养分的土壤异质性会导致不同植被类型的植物组成形成差异[18],RDA分析表明土壤含盐率及土壤含水量可能是影响物种多样性形成差异的重要原因。刘旻霞等[19]研究表明,物种丰富度会随土壤含水量增加而增加。海岸丘陵灌丛在有机质含量较高、水分含量相对较高、水土保持能力较好的海岸丘陵上,因此物种丰富度、多样性最高,这与程浚洋[20]对东部海岛植物的研究结果基本一致。相反,砂生草甸土壤较为瘠薄,含水量较低,含盐量及有机质含量为0.08‰~1.53‰、0.39‰~1.80‰[21],物种丰富度最低。因此土壤异质性可能是导致丘陵灌丛与砂生草甸的物种多样性差异显著、物种相似度极低的原因。另一方面,基岩灌丛与基岩草丛主要分布在土层较薄,土壤性质为风化砂砾土的基岩海岸的石头缝中,土壤养分及含水量相似[14],但基岩草丛分布区域地势较低,更容易受到人为干扰,故基岩草丛丰富度高于基岩灌丛,符合“中度干扰假说”即在一定程度的干扰下,物种丰富度会随干扰程度增加而增加[22]。
4.1.2 不同植被类型的功能多样性
功能丰富度反映物种性状所占据的功能空间大小,功能均匀度反映物种功能性状的空间分布均匀度,体现物种对资源的利用率[23],丘陵灌丛的功能丰富度、功能均匀度显著高于其他3个植被类型,说明丘陵灌丛中的物种功能性状特征丰富,生态位重叠效应弱[24],有效资源利用率高;而砂生草甸的功能丰富度、功能均匀度较低,其生态位空间存在较大空白,有效资源也未得到充分利用,易受到外来入侵植物的侵占。砂生草甸多见于沙滩地,主要由老鼠艻(Spinifex littoreus)、海边月见草、狗牙根(Cynodon dactylon)等喜砂生耐砂生耐旱耐贫瘠植物组成,因此物种功能性状较为单一,功能多样性指数均为最低,物种生存策略差异小而导致物种共存能力下降,功能冗余减少,容易导致生态系统在受到环境动荡时的反应能力及修复能力不足,与丘陵灌丛形成显著差异。功能离散度反映物种间性状功能特征分布位置的差异程度[25],基岩灌丛的Rao指数最高,群落内功能性状差异较大,性状分布较分散,资源竞争小;而基岩草丛功能离散度低于基岩灌丛,或与物种数量的减少所导致的功能性状丢失有关,其生产力及生态系统稳定性不如基岩灌丛。与多数研究结果一致,功能性状差异越大,功能多样性指数越高,生态系统中的有效资源利用率越高,生态系统抵抗非生物因子的功能越强,愈加稳定[5,25-26]。
4.1.3 物种多样性与功能多样性及环境因子的关系
前人对物种多样性和功能多样性的关系一直存在争议[23]。本研究中,功能丰富度与物种丰富度呈极显著正相关,和大多数研究结果一致[12,27],群落中物种数越多,物种功能特征的变化幅度必定越大[28],生态位空间不断分化,功能冗余增加,功能丰富度也就增加。功能均匀度与物种丰富度指数呈极显著正相关,与Pielou均匀度指数呈显著正相关,与Simpson优势度指数呈显著负相关。森林木本群落[9]、森林草本群落[29]、高寒草甸生态系统[10]均发现功能均匀度与物种丰富度无显著关联。一般来说,物种均匀度越高,植物性状集中程度越高,功能离散度越低,资源竞争就越强[27],而本研究同邵丹[30]在云蒙山国家森林公园的研究一致,认为Rao与Pielou均匀度呈极显著正相关;而郑丽婷等[12]通过对庙岛典型植物的研究却发现Rao指数与Pielou均匀度指数没有显著相关性。向琳等[27]在井冈山鹿角杜鹃(Rhododendron latoucheae)群落中发现Rao指数与Simpson优势度指数显著相关,且认为优势度高的群落,功能离散度较小;非随机过程的生态位理论认为,物种在特定环境的限制条件下,表型性状上会出现相似性,物种功能性状会出现聚集性[31],而本研究中海岛植被在恶劣环境胁迫下,功能性状特征存在趋一性[32],其灌草木质化及肉质化程度高、叶较小、叶面被毛,即使优势度高的群落其性状特征分布均匀,因此优势度与功能丰富度及离散度无显著关联。植物多样性往往受到环境因子的影响,在研究区域尺度较小时,尤其是在生态系统敏感、相对独立的海岛,环境因子的作用异常明显[12]。物种多样性往往受到土壤含水量、含盐率、土壤容重等土壤因子的影响[12,29,33],因此在土壤条件较好的丘陵灌丛,其物种多样性与功能多样性相对较高,因此拥有相对稳定的生态系统及较高的生产力。研究表明坡向会影响相关环境因子进而影响物种的分布格局、对资源的利用程度[26-27],尤其是在海岛,坡向能改变风速和风向,还能影响土壤水分,而海拔对环境水热条件至关重要[33]。因此坡向和海拔是影响物种多样性与功能多样性的重要因子,而功能多样性还会受到坡度的影响[19],土壤含水量及土壤容重严重限制着功能丰富度及功能均匀度,制约物种性状进一步分化。
4.2 结论
本研究揭示了海岸不同植被类型的物种多样性和功能多样性特征及两者间的关系。结果表明4种植被类型的物种相似度低;丘陵灌丛的物种丰富度、Shannon-Weiner多样性指数、功能丰富度、功能均匀度高于其他植被类型,砂生草甸的Simpson优势度指数最高;功能丰富度与物种丰富度、Shannon-Weiner多样性指数呈极显著正相关,功能均匀度与4个物种多样性指数均呈显著相关,功能分散度与Shannon-Weiner多样性指数、Pielou均匀度指数显著正相关。土壤容重和坡向是物种丰富度和Shannon-Winner多样性指数、功能丰富度、功能均匀度的主要影响因子,土壤含水率和土壤含盐量是Pielou均匀度指数、Simpson优势度指数的主要影响因子,而功能离散度受土壤含水量、海拔影响较大。综上所述,丘陵灌丛的生态系统较为稳定,基岩灌草丛次之,砂生草甸生态系统较为脆弱,环境因子对物种多样性及功能多样性有着显著的影响,其中土壤水分含量、含盐量、坡度及坡向是主要因子。
猜你喜欢
中国交通信息化(2022年7期)2022-10-27
科学技术创新(2022年30期)2022-10-21
课外生活·趣知识(2021年2期)2021-05-24
学校教育研究(2021年24期)2021-03-28
湖北畜牧兽医(2017年4期)2017-05-18
绿色科技(2017年7期)2017-05-12
科技资讯(2014年8期)2014-11-10
纺织导报(2014年8期)2014-10-31
中国三峡建设(1999年7期)1999-04-06
