
利用SSR标记构建板栗初级核心种质
2021-12-24 10:52张馨方张树航王广鹏
中国农业大学学报 2021年12期
张馨方 张树航 李 颖 郭 燕 王广鹏
(河北省农林科学院 昌黎果树研究所,河北 昌黎 066600)
植物种质资源是开展遗传研究和育种工作的关键,世界各国都高度重视种质资源的搜集、保存和研究利用。随着搜集到的种质数量日益增多,如何对数量庞大的资源进行整理保存,如何高效利用现有种质资源并实现可持续发展,是资源管理单位和育种工作者共同面临的现实问题。鉴于此,Frankel等[1]最早提出核心种质的概念,即以最小的资源数量和遗传重复最大限度地代表整个种质资源的多样性。核心种质是进行优良基因挖掘和资源深入研究的核心子集,能够有效提高资源的利用率。据报道,目前包括桃(Prunuspersica(L.) Batsch.)[2]、核桃(JuglansregiaL.)[3]、木荷(Schimasuperba)[4]和橄榄(Canariumalbum)[5]等木本植物均已构建了核心种质。
中国是栗属(Castanea)植物的起源中心,也是世界上栗属植物分布最为广泛的区域,主要分布有板栗(C.mollissimaBl.)、锥栗(C.henryi(Skam) Rehd. et Wils.)、茅栗(C.seguiniiDode)和日本栗(C.CrenataSieb. & Zucc)4个种[6]。其中板栗在我国分布最广且数量最多,遗传多样性也最为丰富,历来是世界栗属植物遗传改良和新品种培育重要的基因资源[6]。板栗属多年生木本植物,树体高大,资源保存通常以田间活体种植为主,占地面积大,管理费用高。如何从丰富的板栗资源中快速而准确地挖掘出育种工作所需要的优异基因是板栗遗传育种亟需解决的重要问题。据此,本研究启动了构建板栗初级核心种质的策略,优先对核心种质进行繁殖、交换、评价和利用,缓解基因库中庞大资源数量与高效保存之间的矛盾,推进优异资源利用和优良基因挖掘,提高种质资源库的管理和利用水平。
国内外学者对核心种质的构建方法研究较多,包括基于形态学指标构建核心种质[7-8]、基于随机扩增多态性DNA标记(RAPD)数据进行标记辅助取样(Marker-assisted sampling)[9]和利用遗传距离取样等方法[10]。表型性状是多种因素综合作用的结果,易受外界环境影响,加之板栗是高大的多年生乔木,要获得可靠的表型性状数据需要耗费较长年限。而DNA分子标记技术具有高效、快速和稳定等优点,一般较少受到外界环境影响,普遍认为是评价植物遗传多样性和构建核心种质的有效工具。RAPD、扩增片段长度多态性(AFLP)和简单重复序列(SSR)等技术[11-13]已被广泛应用于栗属植物遗传多样性评价方面[14],Marshall和Brown[15]认为位点的等位基因数是评价遗传多样性最重要的指标。程丽莉[16]利用AFLP分子标记对燕山地区136份实生板栗进行了遗传多样性分析,并初选核心种质37份;马玉敏[17]以板栗野生株系为材料,研究了采用荧光AFLP分子标记构建中国野生板栗核心种质的方法;Pereira-Lorenzo等[18]利用SSR分子标记建立了271份欧洲栗(C.sativaMill.)品种的数据信息并筛选出37份核心种质。但有关中国板栗核心种质构建的研究还有待进一步完善,前期研究所选用的材料样本取样量小且取样范围相对狭窄,没有涵盖华北、长江中下游、西北、东南和西南5个地方品种群,遗传信息不够丰富。SSR标记是共显性遗传,能直接反映出DNA序列水平上的变化,并提供丰富的等位基因信息,较其他标记具有更大优势[19]。本研究利用SSR标记,以品种群分布范围涵盖5个地方的279份板栗资源为材料,利用Simple Matching(SM)遗传相似系数和Dice遗传相似系数进行非加权算术平均聚类法(UPGMA)聚类,比较位点优先取样法和随机取样法的不同,确定构建板栗初级核心种质的适宜方法,并筛选出板栗初级核心种质,以期为板栗种质资源的管理、保存和开发利用提供科学依据,有效提高整个行业资源利用率和育种效率。
1 材料与方法
1.1 试验材料
资源植株全部定植于河北省农林科学院昌黎果树研究所板栗种质资源圃(119°15′ E,39°72′ N),属温带半湿润大陆性气候区,年平均气温11 ℃,年平均降水量638 mm,无霜期186 d。所有资源是在中国板栗5个地方品种群分布范围内选取的实生良种、地方品种或古树资源,2004年统一嫁接入圃保存。供试材料为来自河北、山东、北京、陕西、湖南、湖北、江苏、浙江、安徽、贵州、广西和广东12个省(市或自治区) 的279份板栗种质。2018年5月,每个植株选取嫩叶5 ~ 6片,置入装有硅胶的塑封袋带回实验室,置于-80 ℃冰箱中保存备用。供试材料名称及来源地见表1。
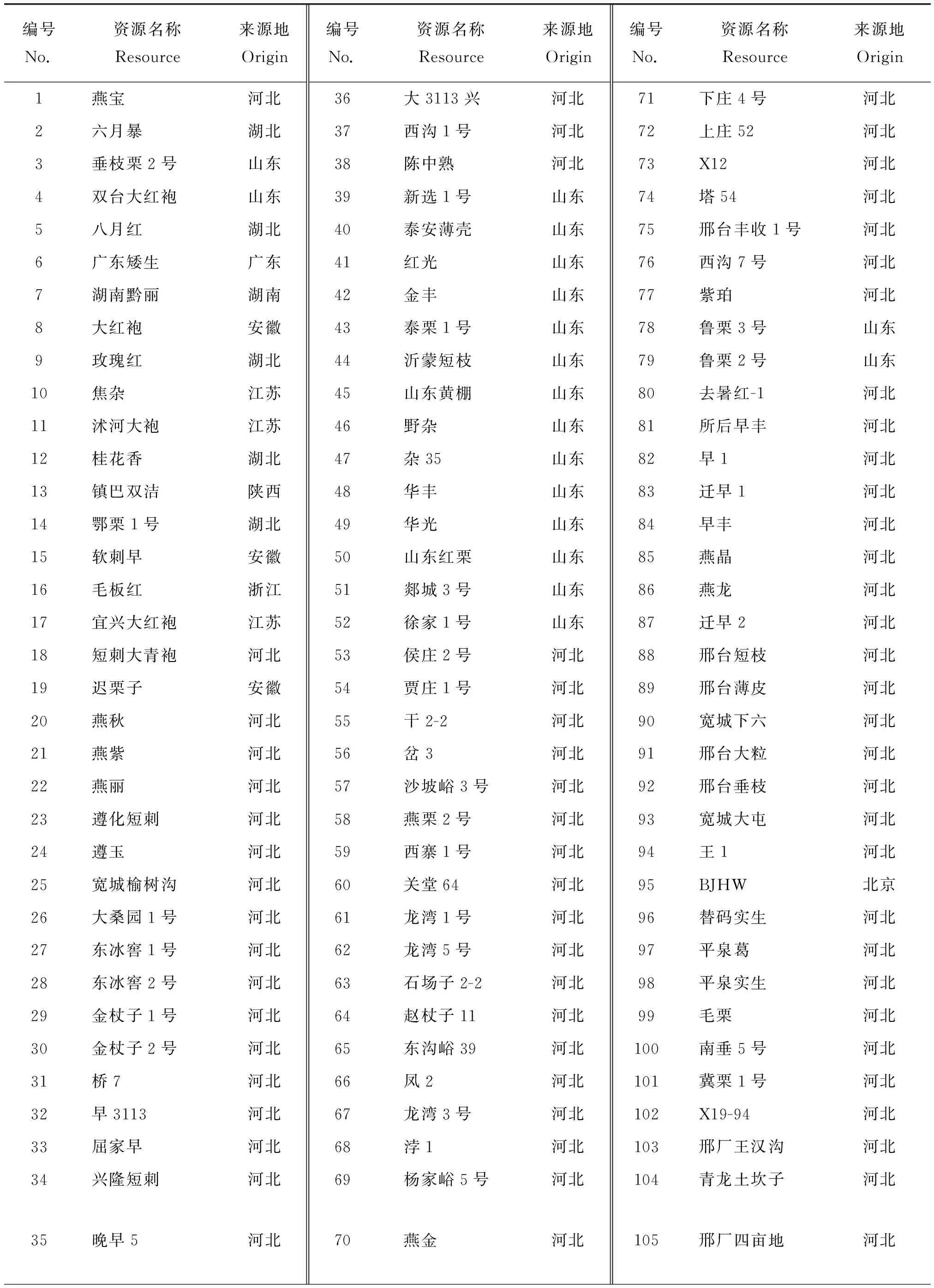
表1 供试板栗资源名称及其来源Table 1 Chestnut materials used in the experiment and their origins
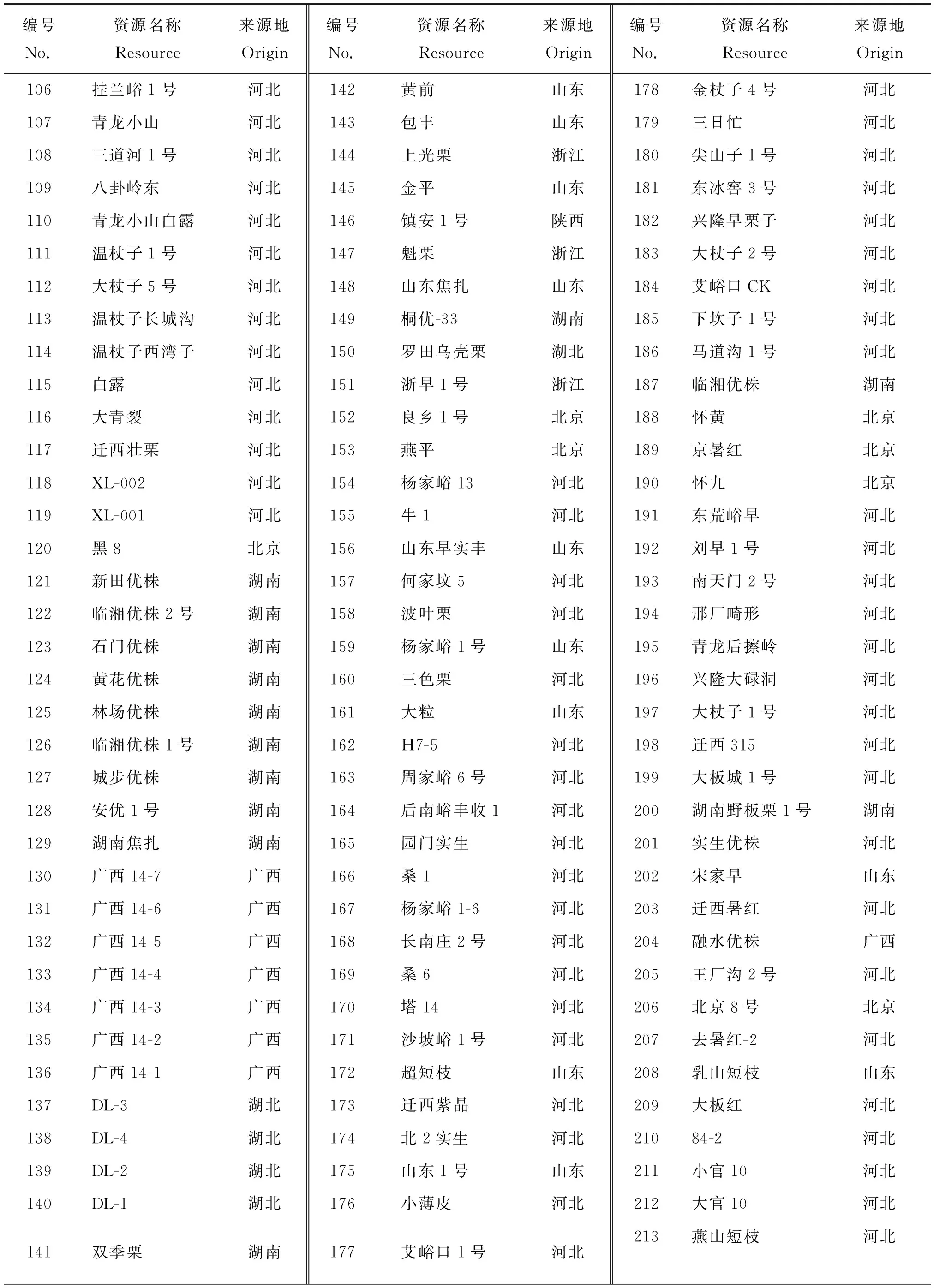
表1(续)

表1(续)
1.2 SSR分析
采用改良的CTAB法[20]提取板栗叶片基因组DNA,用1%琼脂糖凝胶电泳检测DNA浓度,于-20 ℃ 保存备用。本研究从220对引物中筛选出扩增产物多态性较高且条带清晰的21对引物用于全部样品分析,引物具体信息、PCR反应体系及扩增条件参见文献[21]。用6%聚丙烯酰胺凝胶电泳分离扩增产物,用银染法[22]染色。根据DNA marker大小,按照引物设计目标产物条带读取序列长度,在相同迁移位置上,有条带记为“1”,无条带记为“0”。
1.3 初级核心种质构建
利用NTSYS Version 2.10软件,根据SM遗传相似系数和Dice相似系数采用UPGMA法对扩增结果进行聚类分析。SM系数和Dice系数是一种相似性配对系数,即相似匹配数与总匹配数的比值[23],具体计算方法见表2。

表2 遗传相似系数计算方法Table 2 Calculation methods of two genetic similarity coefficients
本研究采取多次聚类的方法,以随机取样法[26]为对照,应用位点优先取样策略[27]构建初级核心种质。位点优先取样策略即根据聚类树状图,在最低分类水平的2份遗传材料中选取稀有等位基因数较多的材料进入下一轮聚类。若2份材料稀有等位基因数目相等,则优先选择稀有等位基因频率值更小的材料,如果这2个值仍相等则随机选择1份材料,若组内只有1份材料则直接进入下一轮聚类。随机取样法即在最低分类水平的2份材料中随机选取1份进入下一轮聚类。
1.4 初级核心种质有效性分析检验
对原种质、保留种质和初级核心种质的有效等位基因数(Ne)、Nei’s遗传多样性指数(H)和Shannon’s信息指数(I)进行t检验,以此评价初级核心种质的代表性。Ne是反映群体遗传变异程度的指标,与等位基因频率有关,而H和I都是反映群体遗传多样性水平的重要参数。以上数据采用POPGENE Version 1.32和SPSS 20.0软件进行分析。
1.5 主坐标分析与表型特征确认
基于SSR数据信息,利用NTSYS Version 2.10软件中的EIGEN模块对初级核心种质的代表性进行主坐标分析,判定初级核心种质是否在主坐标图中均匀分布。
初级核心种质基本形态学指标检测按照《板栗种质资源描述规范和数据标准》[28]进行,从树姿、枝干颜色、皮孔大小和皮孔密度、叶片颜色和形状、叶缘形状、花芽形态和大小以及边果形状方面对初级核心种质进行评价,以此评价初级核心种质的代表性。树姿包括直立、半开张、开张和披垂;枝干颜色包括红褐、灰褐和绿褐;皮孔大小和花芽大小包括小、中和大3种类型;皮孔密度包括稀、中和密;叶片颜色分为浓绿、灰绿、黄绿和紫红色;叶片形状分为椭圆形、阔披针形和披针形;叶缘形状分为锯齿形和钝齿形;花芽形态包括扁圆形和圆形;边果形状包括椭圆形、圆形和三角形。
使用数显游标卡尺(精确度为0.01 mm)和电子天平(精确度为0.01 g)对原种质和初级核心种质的坚果纵径、坚果横径、坚果厚度、单粒质量、果壳质量和果壳厚度6个性状进行测定。参照Hu等[26]评价方法,若均值差异百分率(MD)<20%,同时极差符合率(CR)>80%,可认为核心种质能代表原种质资源的遗传多样性。
(1)
式中:n为数量性状总数;St为核心种质与原种质t测验得到的均值差异显著的性状数。
(2)
式中:n为数量性状总数;RC(i)为核心种质某个性状的极差;RI(i)为原种质同个性状的极差。
2 结果与分析
2.1 初级核心种质构建
由表3可知,除根据Dice系数第1轮聚类后随机取样法取得的样本Ne值略高于位点优先取样法,其他无论利用SM系数还是Dice系数聚类,均为位点优先取样法得到的种质Ne、H和I高于随机取样法。因此,构建板栗初级核心种质时位点优先取样法要优于随机取样法。此外,随着聚类次数的增加,聚类抽样得到的样本数越来越少,占原种质比例越来越低,而Ne、H和I却随聚类次数的增加越来越大。
应用位点优先取样法,279个样本使用SM系数经过3次聚类抽样后(表3),得到了由68份种质组成的核心样本群SA3,占原种质的24.37%,Ne、H和I分别为1.539、0.329和0.502;利用Dice系数聚类,全部样本经3次聚类,得到一个由31份种质组成的核心样本群DA3,占原种质的11.11%,Ne、H和I分别为1.562、0.341和0.517。
2个核心样本群SA3和DA3遗传多样性各指标与原种质的t检验结果见表4。SA3和DA3样本群的Ne、H和I在概率0.05水平上显著大于原种质,说明筛选出的2个核心样本群遗传多样性较丰富。从资源来源地看,样本群DA3包括的种质数量较少,地理来源较单一;而样本群SA3种质来源更丰富,包括原资源来源地10个省(市或自治区),由此初选种质SA3为板栗核心种质。综上,采用位点优先取样法,利用SM相似系数进行3次聚类选取的68份种质,范围涵盖了原材料各主要来源地,经检验能够代表279份原种质的遗传多样性,确定为板栗初级核心种质的最佳取样策略。
2.2 初级核心种质有效性分析检验
保留种质指从原种质中去除核心种质后剩余的部分,它可以作为核心种质的后备资源。对初级核心种质与原种质和保留种质的遗传多样性进行评价,以此判定初级核心种质代表性。初级核心种质和保留种质遗传多样性各指标见表5。结果显示,初级核心种质保留了原种质24.37%的样品,Ne、H和I分别为1.539、0.329和0.502,均高于原种质各遗传多样性指标。由此可见,本研究筛选出的初级核心种质在遗传多样性上能较好地代表原种质。保留种质保留了原种质75.63%的样品,Ne、H和I分别为1.504、0.305、0.468。再对初级核心种质和保留种质的Ne、H和I值分别作t检验(表4),初级核心种质Ne、H和I在概率0.05水平上显著大于保留种质,因此,应优先考虑应用初级核心种质资源进行遗传研究工作。
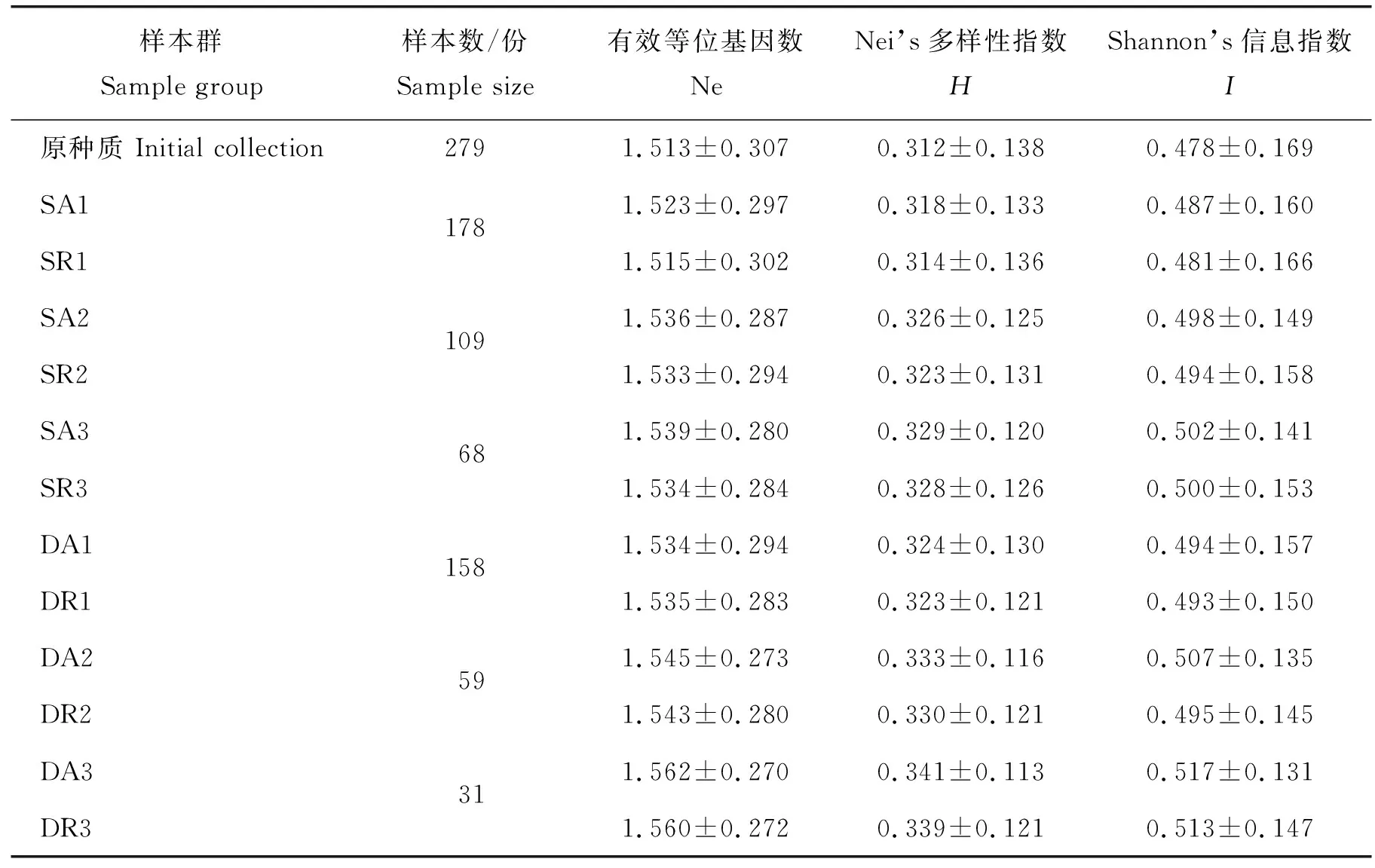
表3 板栗不同样本群遗传多样性比较Table 3 Genetic diversity analysis of different collections of China chestnut
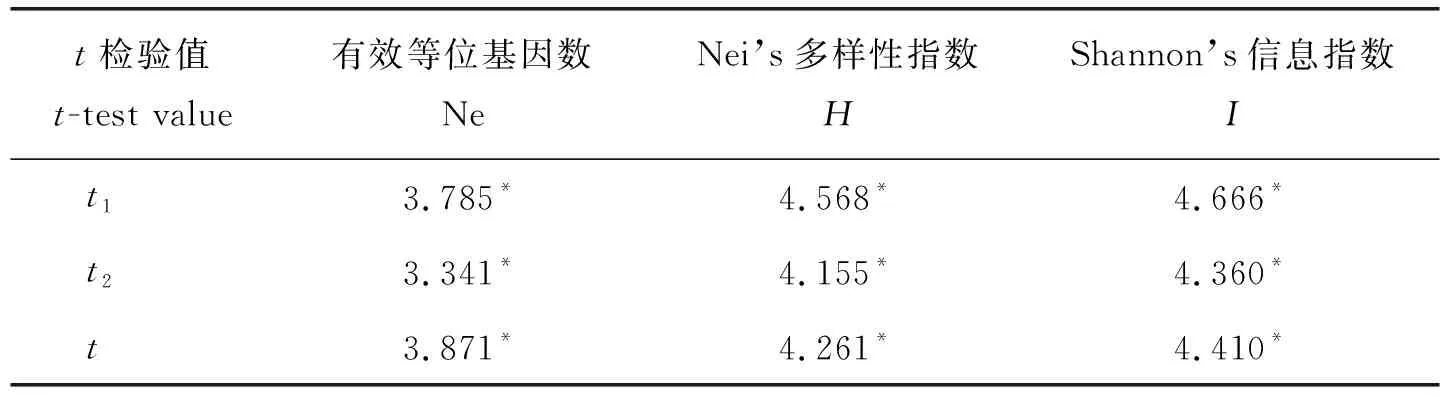
表4 2个核心样本群与原种质、初级核心种质与保留种质遗传多样性参数的t检验Table 4 Gentic diversity comparison and t-test between two core sample group and initial collection, primary core colletion and reserved collection
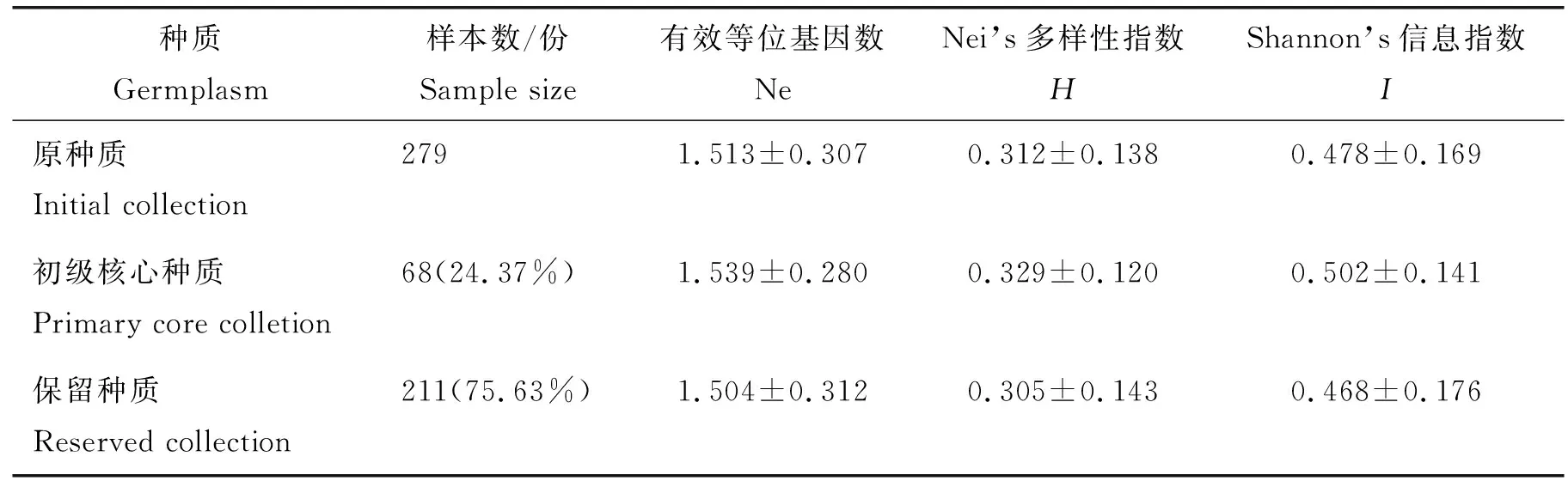
表5 初级核心种质和保留种质、原种质遗传多样性对比Table 5 Genetic diversity comparison and t-test between core collection, reserved collection and initial collection
2.3 初级核心种质主坐标分析与表型特征评价
采用主坐标法(PCoA)对筛选出的初级核心种质进行确认,绘制原种质与初级核心种质1、2主坐标散点图(图1),可见初级核心种质在整个板栗资源的主坐标图分布均匀,表明筛选出的初级核心种质具有较好的代表性。
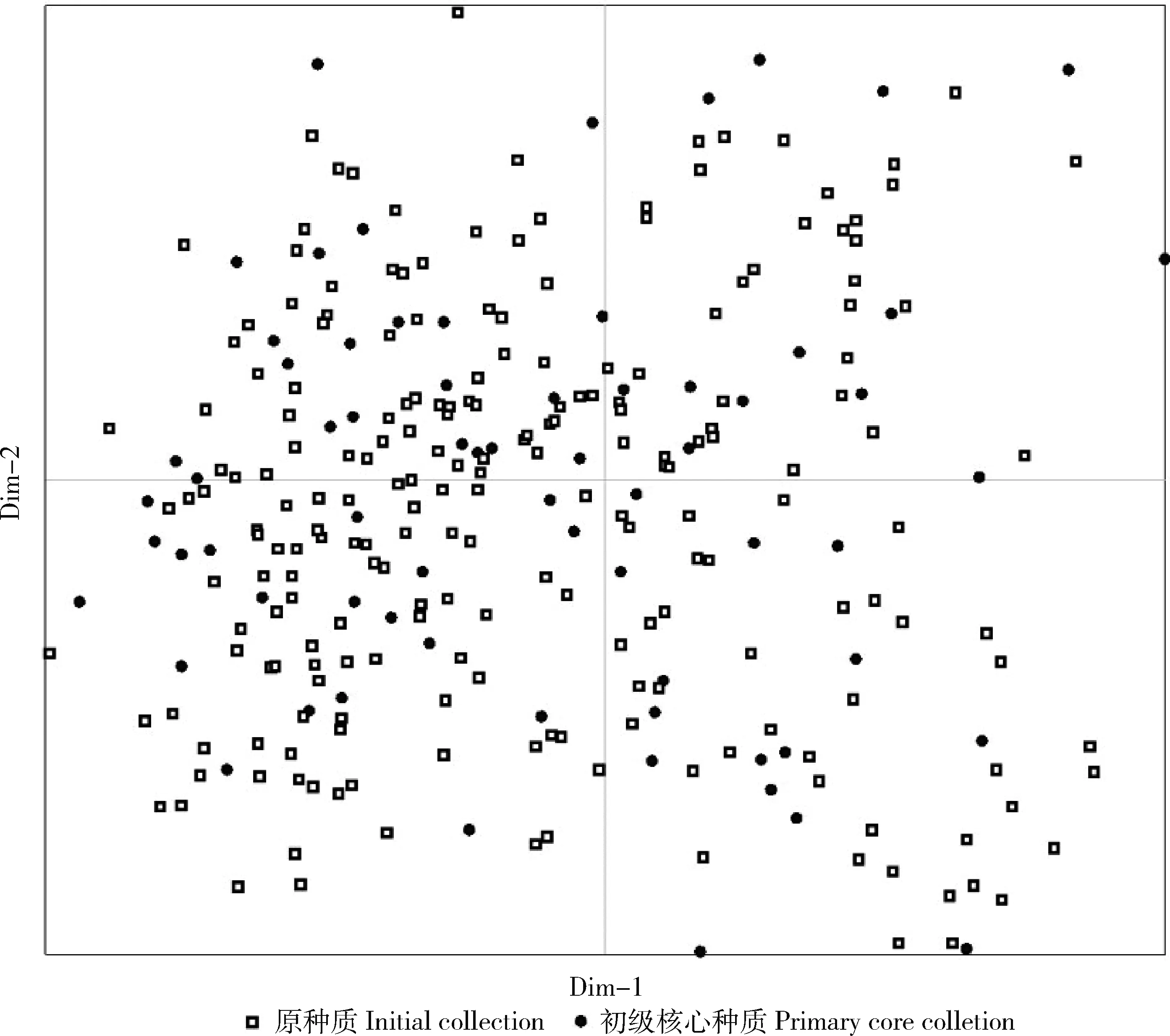
横纵坐标分别表示贡献率较高的1、2主坐标。 Horizontal and vertical coordinates mean the first two principal coordinates with higher contribution rate respectively.图1 位点优先取样法构建的初级核心种质与原种质的主坐标图Fig.1 Principal coordinates plots of primary core collection constructed by preferred sampling strategy and initial collection
本研究构建的68份初级核心种质的编号分别为1、2、8、10、12、15、17、24、26、40、44、46、50、51、53、54、57、59、65、66、76、92、94、98、101、116、118、123、125、128、147、149、151、158、160、161、166、173、174、177、180、183、185、186、191、203、209、214、215、222、223、231、232、233、238、242、244、254、255、256、260、263、264、269、270、271、275和277,68份初级核心种质名称及来源地见表1。68份种质在树姿、枝干颜色、皮孔大小、皮孔密度、叶片颜色和形状、叶缘形状、花芽形态和大小以及边果形状10个方面涵盖了分类学的大多数或全部类型。此外还包括红栗、垂枝栗、三色栗、波叶栗和短枝型板栗等具有特殊性状的资源,验证了本研究筛选出的初级核心种质的合理性。
初级核心种质与原种质6个表型特征数据见表6。经t检验核心种质6个表型性状均值与原种质无显著差异,根据公式计算出MD和CR分别为0和81.13%,符合胡晋等[29]提出的核心种质均值同原种质群体存在显著差异的性状<20%、核心种质与原种质的极差符合率>80%的最低标准。研究结果从表型水平上证明了根据SM系数聚类结合位点优先取样法是构建板栗初级核心种质较适宜的方法。
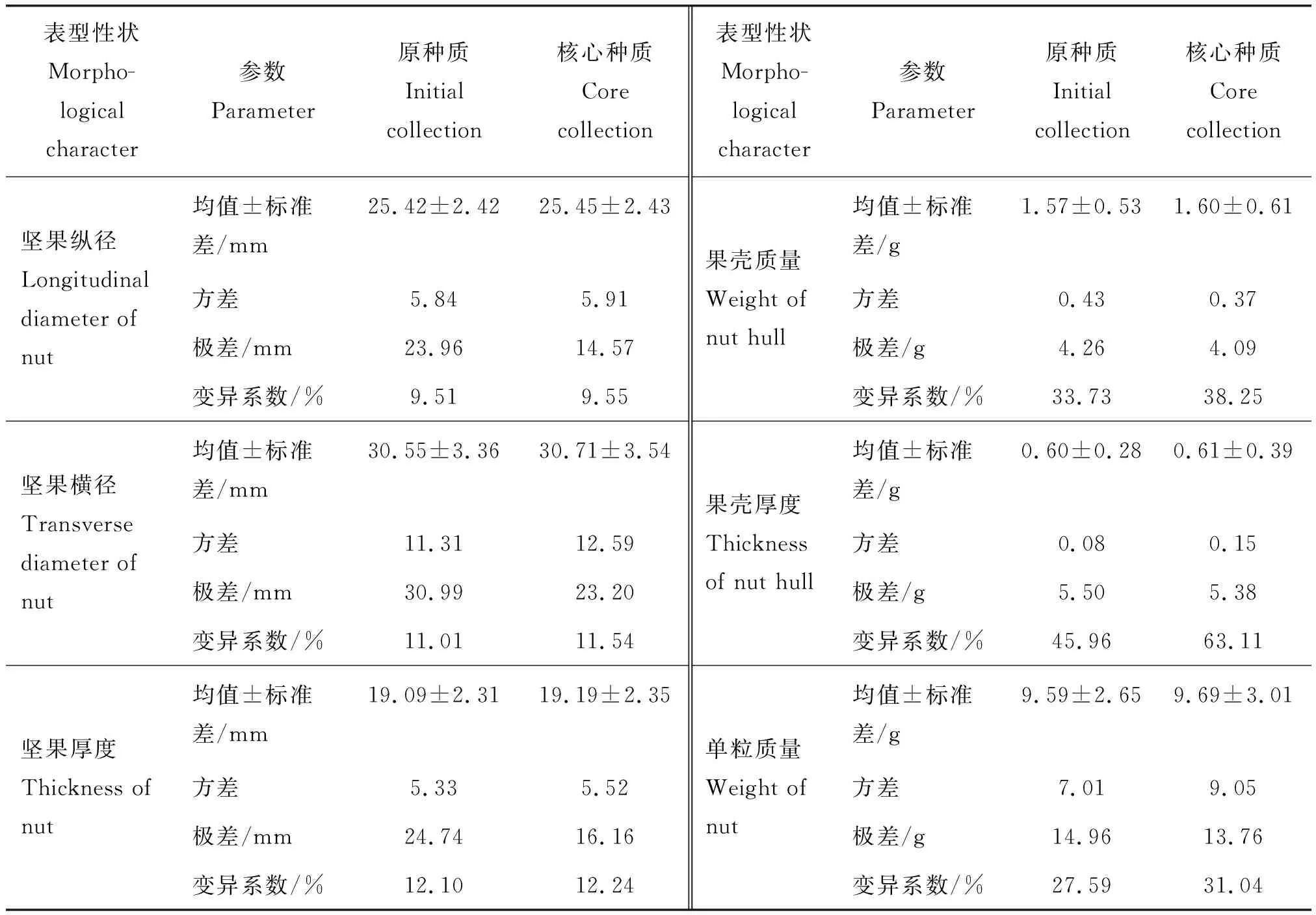
表6 初级核心种质与原种质6个坚果表型特征基本参数Table 6 Basic parameter value of 6 phenotypic traits of primary core collection and initial collection
3 讨论与结论
在明确资源分子遗传信息数据的基础上,本研究应用聚类分析的方法构建核心种质,因为具有最大遗传相似度或最近遗传距离的材料自然聚在一起,而去除遗传距离较近的材料后可得到遗传特性差异较大的种质材料。目前,基于遗传距离聚类的取样方法已经建立并应用于植物核心种质的构建当中[10,30-31]。研究表明,选择不同遗传相似系数导致分析结果存在较大差异,因为遗传系数的选择会影响遗传相似度或遗传距离的计算,因此,选择合适的遗传相似系数对于准确估计个体间遗传相似度和评价群体间遗传多样性至关重要[23]。刘娟等[32]研究认为,构建新疆野杏核心种质时采用Dice遗传距离要优于SM遗传距离;张春雨等[27]认为,根据SM、Jaccard或Dice遗传距离进行多次聚类构建新疆野苹果核心种质均为较适宜的方法,并提出了对于共显性分子标记如SSR和限制性内切酶片段长度多态性(RFLP),利用Dice和Jaccard相似系数更为理想。本研究采用SM遗传系数和Dice系数通过多次聚类的方法构建板栗初级核心种质,结果表明利用Dice系数取得的核心样本数量最小,却具有较大的Ne、H和I,说明从遗传系数的选择而言,Dice系数对SSR标记更合适,符合张春雨等研究结果。但考虑到采用Dice系数构建的核心样本群地理来源范围较单一,未能包括所有样本来源地,而利用SM系数构建的核心样本群来源地涵盖原种质10个省(市),在遗传多样性上能够以最小的样本量最大程度地代表原种质,因此本研究优先采用SM遗传相似系数进行聚类分析。
取样策略是核心种质构建中又一重要环节。李自超等[33]通过研究水稻核心种质的构建方法发现聚类法要优于随机法。通常不能采用在整个群体内完全随机的方法进行取样,因为一个物种的遗传多样性是以某种特定组织和结构分布,而不是完全随机地分布在整个群体中[34-35]。Quero-Garcia等[36]在构建芋头核心种质研究中,比较了完全随机法取样、组内完全随机法取样和组内先聚类后取样的方法,结果表明组内先聚类后取样是一种比前两者更加有效的方法。本文即采用先聚类后取样的方法,研究表明无论选择何种遗传系数聚类,通过位点优先策略取得的样本均比随机取样法具有较大的Ne、H和I,这说明构建板栗初级核心种质时位点优先取样法优于随机取样法,与前人的研究结果一致[27,32]。可能是物种的基因多样性是决定其表型性状的遗传因子,而所有位点的等位基因频率和等位基因数目构成了资源基因多样性的基础。等位基因数目和频率承载着原种质的遗传多样性,而稀有等位基因可能具有某种有利特性来帮助该物种适应不同的环境变化,防止群体衰落,在生物多样性中具有重要意义[37-39]。因此,建议在构建核心种质时优先选取稀有基因(等位基因频率小于5%),同时尽可能地保留原种质等位基因的多样性,这样既可以减少等位基因的丢失,又尽可能保留原种质群体的基因多样性。
样本量的规模是衡量核心种质是否合理有效的重要因素。核心种质是以最小的资源数量最大程度地代表整个资源的遗传多样性,因此确定最优的取样比例十分关键。李自超等[40]提出应根据具体植物的遗传结构和数量规模来确定相应的取样比例。目前国内外构建的核心种质所占比例一般为原种质的5%~30%,其中作物群体一般原种质资源数量较多,选取较小规模的核心样品也能充分保留原种质的遗传特性。如稻类等农作物核心样品一般占总收集品的10%左右[41-42],而园艺植物核心种质取样规模一般在20%左右[32]。取样比例应根据具体树种遗传特性和原种质资源数量和实际工作而定。对遗传多样性丰富和资源数量大的群体可适当减小取样比例。本研究选取68份初级核心种质占原种质的24.37%,来源涵盖了中国板栗5个地方品种群分布范围,能充分代表原种质的遗传多样性。此外,本研究从主坐标和形态学角度对构建的初级核心种质进行确认,发现初级核心种质遍布整个主坐标图,在表型特征方面能代表原群体,通过位点优先取样策略结合SM相似系数多次聚类确定的24.37%的取样比例是合理有效的。建议板栗行业优先对这部分初级核心种质进行繁殖、交换、评价和利用,以促进各地板栗基因交流,提高整个行业资源利用率和育种效率。而对于资源保存单位来说,这部分初级核心种质的确定可以提高整个种质资源库的管理、保存和利用水平。尽管理论上本研究构建的初级核心种质在遗传多样性方面能最大程度地代表整个供试板栗资源,但其他种质资源仍具有重要的保存意义。因为本研究仅是借助21对SSR引物对其遗传多样性进行了初级地挖掘,今后随着对资源评价内容的拓展和评价水平的提高,资源遗传多样性将得到进一步挖掘,从中仍可能筛选出极具利用潜力的基因资源,其他植物对非核心种质的留存亦可参考如此。
本研究所选用的279份资源取材于中国板栗主产区的12个省(市或自治区),涵盖传统分类学上划定的中国板栗5个地方品种群,是迄今为止进行中国板栗核心种质群构建选用材料(样本)数量较多且覆盖面较广的研究。尽管如此,本研究也存在原种质取样不均的问题,其中华北品种群的资源数量相对较多,其他品种群样本相对较少,可能遗漏部分具有潜在利用价值的种质资源,因此本研究选用样本在代表中国板栗资源的全部遗传信息方面仍有欠缺。此外,由于针对资源表型特征的检测数据相对简单,在以后研究中还需要补充和完善质量性状表型保留或数量性状特征值方面数据,以此来进一步验证核心种质的代表性及合理性。
猜你喜欢
公民与法治(2022年12期)2023-01-07
小学生作文(低年级适用)(2022年11期)2022-12-02
四川蚕业(2022年2期)2022-11-19
四川蚕业(2022年2期)2022-11-19
今日农业(2022年13期)2022-09-15
四川蚕业(2020年4期)2020-02-10
创新作文(小学版)(2019年10期)2019-09-25
小学生作文(中高年级适用)(2018年5期)2018-06-11
中国麻业科学(2018年6期)2018-04-09
西南农业学报(2016年5期)2016-05-17
