
武陵石漠化山地八种针阔叶树种树干液流速率日变化特征
2022-01-04 09:16刘振华童方平李光强
湖南林业科技 2021年6期
陈 瑞, 刘振华, 李 贵, 张 翼, 童 琪, 吴 敏, 童方平, 李光强, 孙 良
(1.湖南省林业科学院,湖南 长沙 410004; 2.贵州省植物园,贵州 贵阳 550004; 3.浏阳市浏阳湖国有林场, 湖南 浏阳 410309)
石漠化是在岩溶及其发育的自然背景下形成的,是喀斯特地区土地退化的极端形式[1-2]。石漠化区土层薄,保水能力差,水分成为了限制石漠化区森林植被恢复与重建的关键生态因子[3-5],因此,石漠化区森林植被耗水已成为生态重建过程中水资源管理首要考虑的重要因素[6]。树木树冠的蒸腾量受树干液流量的制约[7],树干液流参数能够反映植物的蒸腾耗水特性[8-9],而热脉冲技术能在树木自然生长状态下测量树干木质部位上升液流动速度及流量,可简便地测定树冠的蒸腾水量[10-11]。对大部分树种耗水特性的研究主要是通过盆栽进行胁迫实验[12-13],或者用TDP技术对树种长期监测[14-16]来研究树种蒸腾耗水及环境因子影响的规律,目前,针对石漠化区树种的耗水日变化规律报道较少。本文利用TDP技术对武陵山地石漠化区8种乔木树种的树干液流日变化及耗水规律进行监测,以期揭示石漠化区林木耗水规律,从而对湖南大面积石漠化地区的林种布局、树种选择等提供参考。
1 研究区概况
湖南省张家界市桑植县西界林场位于109°40′—110°46′E,29°17′—29°48′N,海拔500~1172 m,属中亚热带季风湿润气候区、山地地貌。其年平均气温11.5~15 ℃,无霜期220~260 d,1月平均气温4.7 ℃,7月平均气温27.4 ℃;年平均降雨量1 400~1 600 mm;春、夏、秋、冬季的日照时数分别为276.8、526.6、314.4、179.1 h[17];母岩主要为石灰岩,土壤主要为黄壤和黄棕壤,土层厚度在30~80 cm。
2 研究方法
2.1 样地设置与样树选取
标准样地设置在西界林场石漠化区自然恢复的针阔混交林内,面积为40 m×60 m。对样地内林木进行每木检尺,记录树种及测量胸径、树高、冠幅(EW×NS)、枝下高等,计算林分平均木和径阶平均木的胸径和树高,并在标准地内标出,用于备选目标树。选择的树种主要有黄樟(Cinnamomumporrectum)、马尾松(Pinusmassoniana)、柏木(Cupressusfunebris)、杉木(Cunninghamialanceolata)、麻栎(Quercusacutissima)、刺楸(Kalopanaxseptemlobus)、木荚红豆(Ormosiaxylocarpa)及朴树(Celtissinensis)。
根据混交林调查的实际情况,不同树种按照不同的胸径跨度划分径阶,再根据各树种胸径分布的实际情况进行最终目标树选择。在最终目标树选择时,将调查数据中胸径梯度三等分划分,每个梯度选择1株目标树。按照试验要求,对胸径大于6cm的目标树,要求树干通直圆满、不偏心、不偏冠,测定部位上、下50 cm处无节疤或损伤。最终确定的目标树见表1。
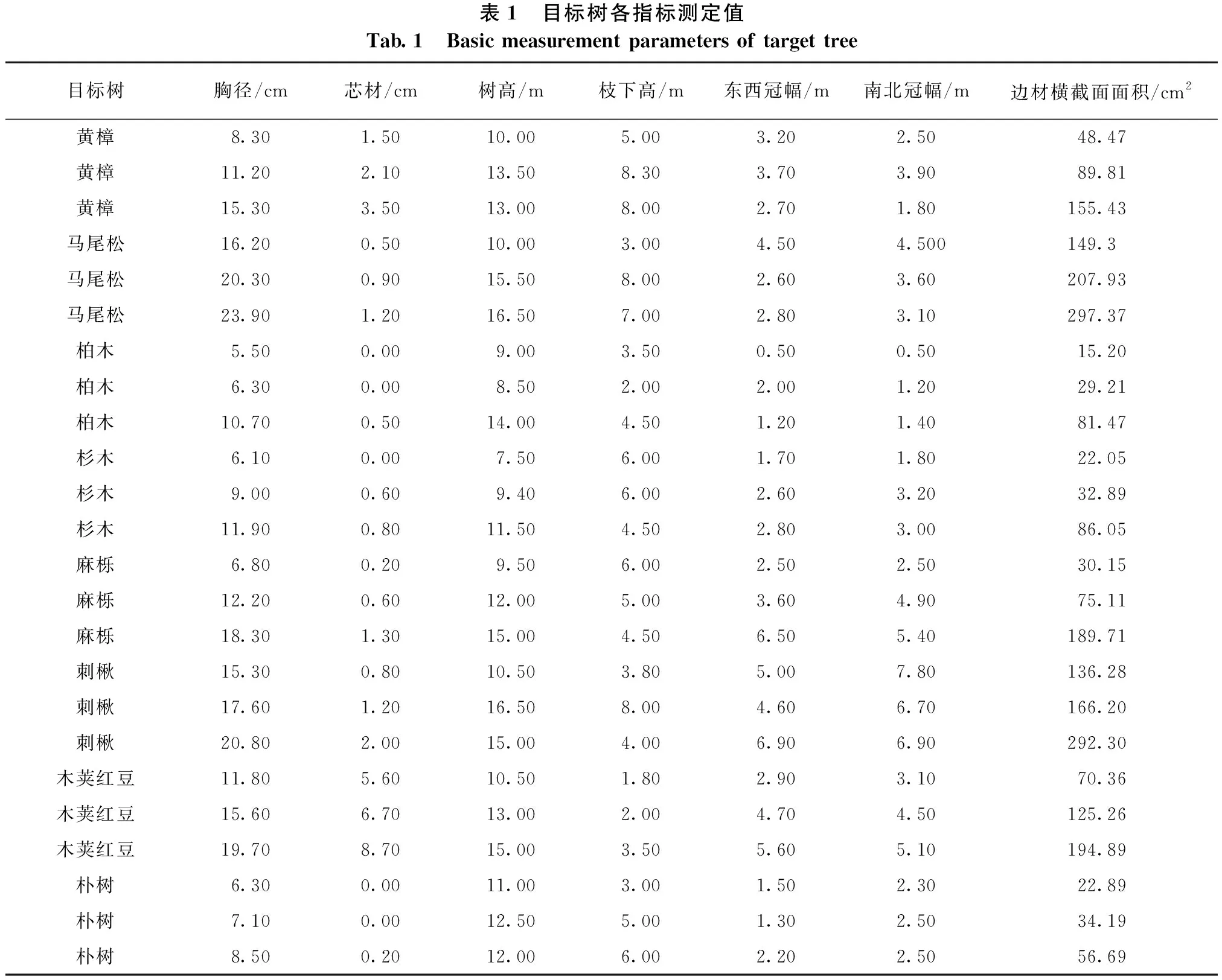
表1 目标树各指标测定值Tab.1 Basic measurement parameters of target tree目标树胸径/cm芯材/cm树高/m枝下高/m东西冠幅/m南北冠幅/m边材横截面面积/cm2黄樟8.301.5010.005.003.202.5048.47黄樟11.202.1013.508.303.703.9089.81黄樟15.303.5013.008.002.701.80155.43马尾松16.200.5010.003.004.504.500149.3马尾松20.300.9015.508.002.603.60207.93马尾松23.901.2016.507.002.803.10297.37柏木5.500.009.003.500.500.5015.20柏木6.300.008.502.002.001.2029.21柏木10.700.5014.004.501.201.4081.47杉木6.100.007.506.001.701.8022.05杉木9.000.609.406.002.603.2032.89杉木11.900.8011.504.502.803.0086.05麻栎6.800.209.506.002.502.5030.15麻栎12.200.6012.005.003.604.9075.11麻栎18.301.3015.004.506.505.40189.71刺楸15.300.8010.503.805.007.80136.28刺楸17.601.2016.508.004.606.70166.20刺楸20.802.0015.004.006.906.90292.30木荚红豆11.805.6010.501.802.903.1070.36木荚红豆15.606.7013.002.004.704.50125.26木荚红豆19.708.7015.003.505.605.10194.89朴树6.300.0011.003.001.502.3022.89朴树7.100.0012.505.001.302.5034.19朴树8.500.2012.006.002.202.5056.69
2.2 树干液流速率测定
2017年7—9月,选择典型晴天,在样树树干离基部1.3 m高处将周围10 cm×10 cm范围内树干外层树皮刮掉,露出内层活树皮;用特定规格钻头在选定部位钻取直径1.8 mm、深80 mm的小孔,插入TDP液流探针;探针的另一端与数据采集器连接,为了防止太阳辐射对探针产生影响,探针安放好后用保温、防辐射的铝箔包住其外部树干,然后用胶带密封。测定样树树干液流时,数据记录间隔为15 min,24 h不间断监测。本试验选择7—9月中旱季和雨季的数据进行分析。
使用Granier公式计算树干液流速率[18]:
Vs=0.011 9K1.231×3 600
(1)
式中:Vs为树干液流速率(g·h-1);K=(dTm-dT)/dT(dTm为树干液流速率为零时的温差最大值,一般在黎明前无径流时测得;dT为瞬时温差值)。
采用生长锥钻取样树木质部来确定样树边材横截面面积,计算公式为[19]:
As=π[(D/2-d)2-r2]
(2)
式中:As为边材横截面面积(cm2);D为胸径(cm);d为树皮厚度(cm);r为心材半径;π值取3.14。
计算边材横截面面积并与V相乘得到体积流量:
Sapflow=A×V
(3)
式中: Sapflow为单株树干液流通量(mL);A为安装液流仪探针处的边材横截面面积(cm2);V为树干液流速率( mL·cm-2)。
2.3 环境因子测定
研究区气象参数通过小型便携式自动气象站来记录,设定与TDP树干液流仪同样的时间间隔(15 min),测定太阳辐射、空气温度、空气湿度、风速等气象因子,数据读取采用Excel 2007。
2.4 数据分析
采用Excel 2007进行做图;采用SPSS 21进行相关性分析。
3 结果与分析
3.1 不同树种树干液流速率日变化特征
各树种树干液流速率平均日变化见图1、图2。

图1 各树种7月树干液流速率平均日变化Fig.1 Average diurnal variation of flow rate of different tree species in July
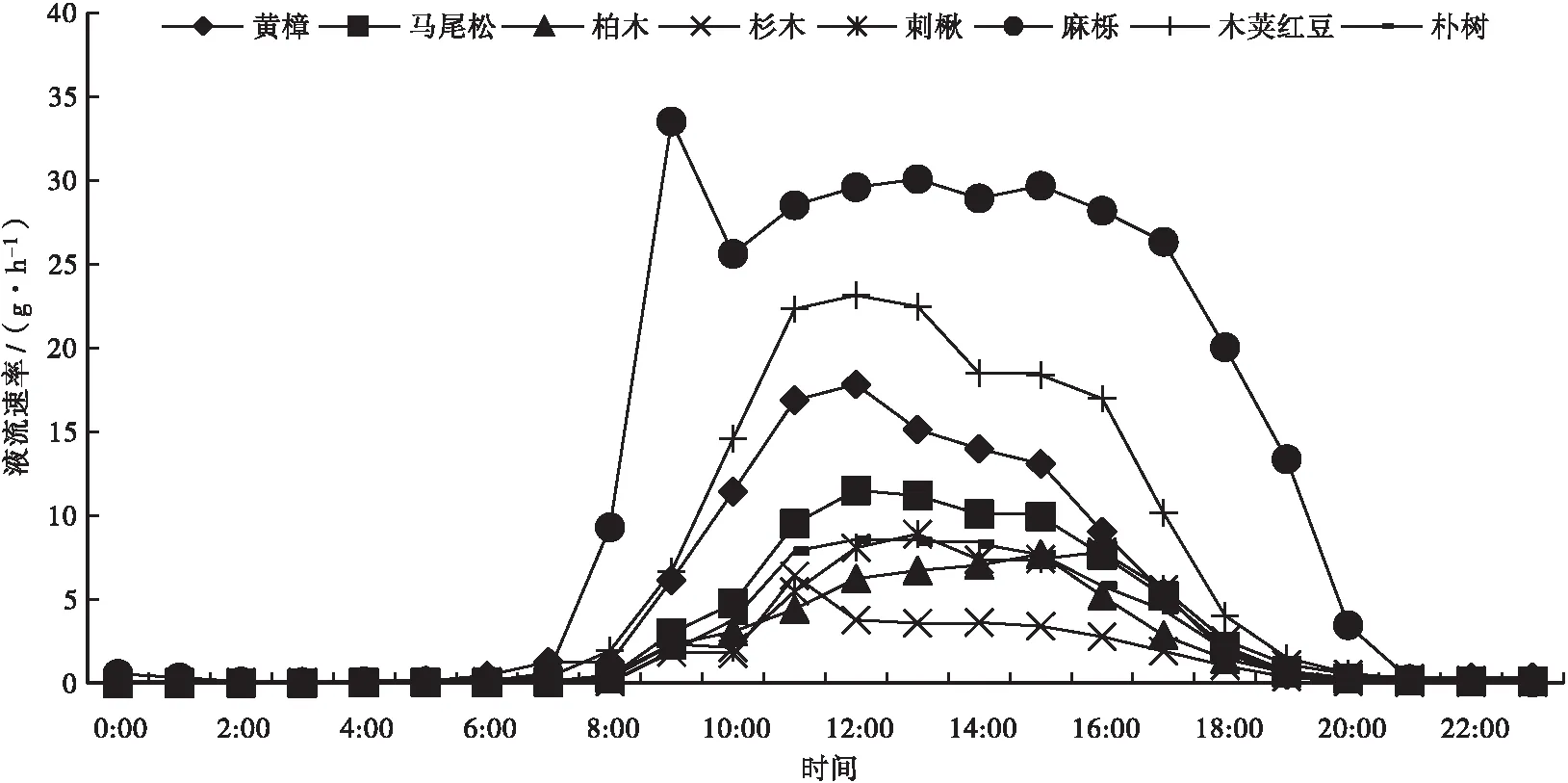
图2 各树种9月树干液流速率平均日变化Fig.2 Average diurnal variation of flow rate of different tree species in September
图1、图2显示:7月和9月8个树种树干液流速率日变化表现出单峰或双峰曲线的趋势,除9月马尾松外,阔叶树的树干液流速率均大于针叶树的。阔叶树的树干液流在7:00—8:00开始启动,针叶树树的干液流在8:00开始启动。8个树种的树干液流速率峰值7月出现在11:00—14:00,9月出现在9:00—15:00;树干液流停止时间7月出现在19:00—22:00,9月出现在19:00—21:00。7月各树种树干液流速率最大值排序为刺楸>木荚红豆>朴树>麻栎>黄樟>马尾松>杉木>柏木;9月各树种树干液流速率最大值排序为麻栎>木荚红豆>黄樟>马尾松>刺楸>朴树>柏木>杉木。
3.2 不同树种平均树干液流通量日变化特征
各树种树干液流通量日变化特征见图3。图3显示:麻栎的树干液流通量日变化呈现出单峰和多峰曲线趋势,7月峰值出现在14:00,峰值为1 116.20 g·cm-2h-1;9月在9:00左右达到一天中的最大值(3 314.31 g·cm-2h-1),随后开始下降,11:00以后出现回升,在13:00到达第2个峰值(2 974.20 g·cm-2h-1)后下降,14:00以后再次出现回升,在15:00到达第3个峰值(2 935.55 g·cm-2h-1)。刺楸的树干液流通量呈现出双峰曲线趋势,7月在11:00左右达到一天中的最大值(3 893.82 g·cm-2h-1),随后开始下降,14:00以后出现回升,在15:00到达第2个峰值(3 363.11 g·cm-2h-1)后下降;9月峰值在13:00左右达到一天中的最大值(1 777.83 g·cm-2h-1),随后开始下降,15:00以后出现回升,在16:00到达第2个峰值(1 566.43 g·cm-2h-1)后下降。木荚红豆树干液流通量呈现出单峰或双峰曲线趋势,7月峰值出现在12:00和15:00,峰值分别是2 394.06 g·cm-2h-1和2 013.62 g·cm-2h-1;9月峰值出现在12:00,为3 088.75 g·cm-2h-1。朴树树干液流通量呈现出单峰或叁峰曲线趋势,7月在9:00左右达到一天中的最大值(445.78 g·cm-2h-1),随后开始下降,10:00以后出现回升,在12:00到达第2个峰值(486.27 g·cm-2h-1)后下降,14:00以后出现回升,在15:00到达第3个峰值(459.71 g·cm-2h-1);9月峰值出现在12:00,为325.26 g·cm-2h-1。其它树种的树干液流通量均呈现单峰曲线趋势,其中7月黄樟和马尾松的树干液流通量峰值均出现在13:00,柏木和杉木的均出现在14:00,黄樟、马尾松、柏木、杉木的分别为1245.65、1783.73、238.00、393.54 g·cm-2h-1。9月杉木的树干液流通量于11:00达到峰值,为314.18 g·cm-2h-1;柏木的树干液流通量峰值出现在15:00,为283.05 g·cm-2h-1;黄樟、马尾松的均出现在12:00,分别为1781.86、1888.15 g·cm-2h-1。
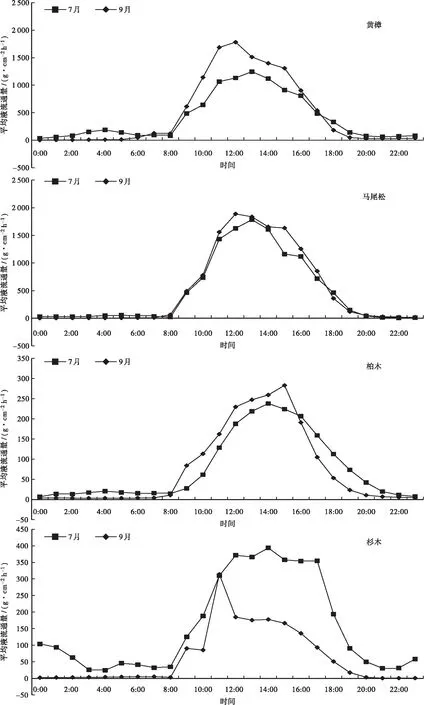
图3 各树种7月和9月树干液流通量日变化特征
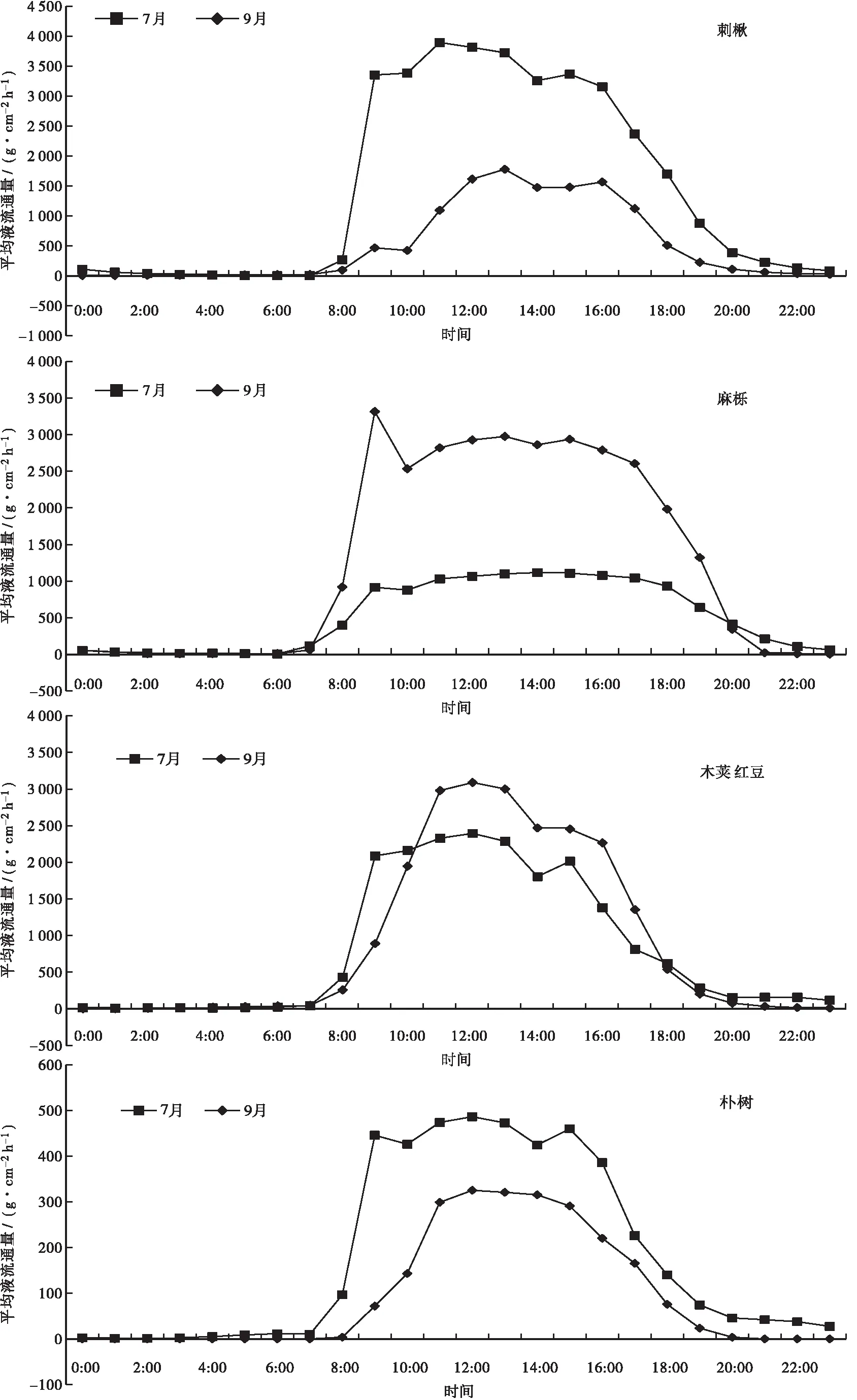
续图3 各树种7月和9月树干液流通量日变化特征
比较8种乔木的树干液流通量日均值,7月的依次为刺楸(1 426.42 g·cm-2h-1)>木荚红豆(805.90 g·cm-2h-1)>麻栎(515.18 g·cm-2h-1)>马尾松(485.95 g·cm-2h-1)>黄樟(399.07 g·cm-2h-1)>朴树(179.58 g·cm-2h-1)>杉木(155.97 g·cm-2h-1)>柏木(77.29 g·cm-2h-1);9月的依次为麻栎(1273.22 g·cm-2h-1)>木荚红豆(906.52 g·cm-2h-1)>马尾松(527.18 g·cm-2h-1)>刺楸(508.12 g·cm-2h-1)>黄樟(482.47 g·cm-2h-1)>朴树(94.21 g·cm-2h-1)>柏木(75.89 g·cm-2h-1)>杉木(63.94 g·cm-2h-1)。表明,阔叶树种的树干液流通量普遍高于针叶树种的。
比较不同树种的树干液流通量,7月刺楸的树干液流通量最大值是柏木的16.36倍,9月麻栎的树干液流通量最大值是柏木的11.71倍。
图3还可以看出:在7:00—15:00,黄樟7月的树干液流通量小于9月的,其它时段的均大于9月的;在8:00—17:00,马尾松7月的树干液流通量小于9月的,其它时段的均大于9月的;在9:00—15:00,柏木7月的树干液流通量小于9月的,其它时段的均大于9月的;在11:00,杉木7月的树干液流通量小于9月的,其它时段的均大于9月的;刺楸、朴树7月的树干液流通量均大于9月的;在8:00—19:00,麻栎7月的树干液流通量小于9月的,其它时段的均大于9月的;在11:00—17:00,木荚红豆7月的树干液流通量小于9月的,其它时段的均大于9月的。从启动时的树干液流通量来看,麻栎、黄樟、柏木7月的小于9月的,杉木、刺楸、朴树7月的大于9月的。
3.3 不同树种平均日耗水量
各树种的平均日耗水量见表2。
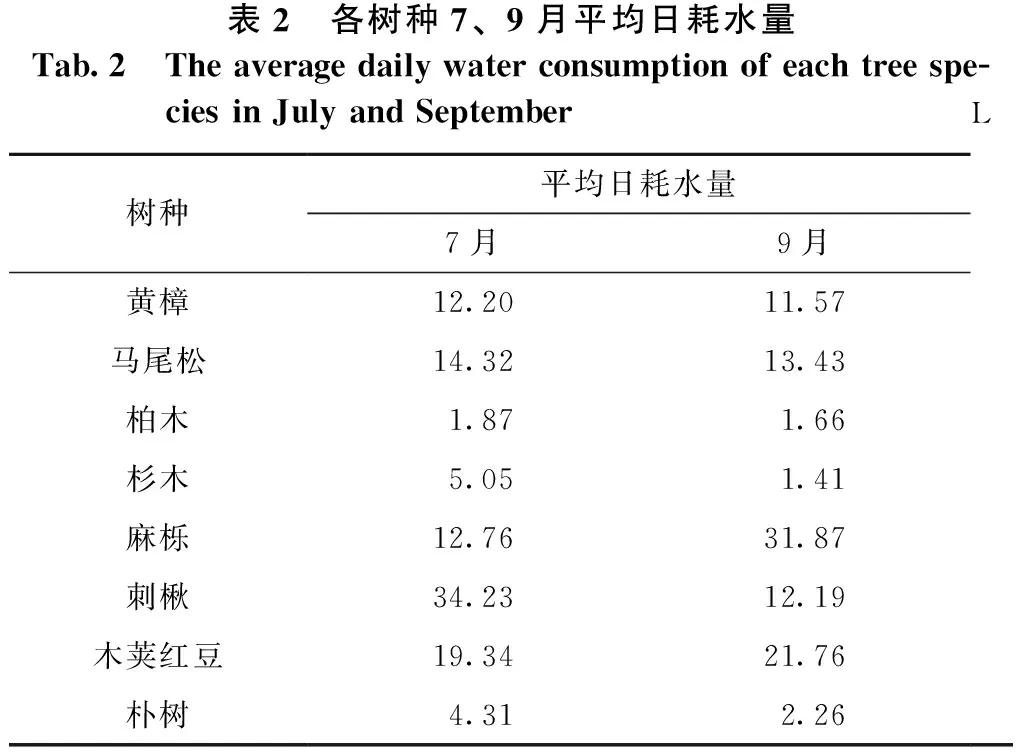
表2 各树种7、9月平均日耗水量Tab.2 The average daily water consumption of each tree spe-cies in July and SeptemberL树种平均日耗水量7月9月黄樟12.2011.57马尾松14.3213.43柏木1.871.66杉木5.051.41麻栎12.7631.87刺楸34.2312.19木荚红豆19.3421.76朴树4.312.26
表2显示:除麻栎、木荚红豆外,其它树种的平均日耗水量都是7月的大于9月的。7月刺楸的平均日耗水量最大,为34.23 L;柏木的最小,为1.87 L;各树种平均日耗水量依次为刺楸>木荚红豆>马尾松>麻栎>黄樟>杉木>朴树>柏木。9月麻栎的平均日耗水量最大,为31.87 L;杉木的最小,为1.41 L,各树种平均日耗水量依次为麻栎>木荚红豆>马尾松>刺楸>黄樟>朴树>柏木>杉木。9月的平均日耗水量相对于7月的减少量最大的是杉木,减少了258.16%;最小的是黄樟,减少了5.45%。
3.4 树干液流速率与气候因素相关性及其模型拟合
从表3可以看出,树干液流速率除了与风速和空气温度无显著相关性外,与其它环境因子均呈显著相关,其中与太阳总辐射的相关性最显著,说明太阳总辐射对树干液流速率的影响比其他环境因子的大。

表3 树干液流速率与气候因子的相关性Tab.3 Correlation analysis of flow rate and climatic factors土壤湿度土壤温度风速空气湿度空气温度大气压强太阳总辐射降雨量树干液流速率-0.036*0.042*0.016-0.035*-0.0290.036*0.047*-0.36* 注:*表示在 0.05 水平(双侧)上显著相关。
表4为各树种树干液流速率与气候因子的多元线性回归模型。从表4看出,在各树种的回归模型中,空气湿度和风速多次被删除,说明树干液流速率与空气温度、土壤温度、土壤湿度、大气压强、太阳总辐射的相关性较强。拟合模型因树种不同而存在一定差异,表明树干液流速率与气候因子之间的关系较复杂。

表4 各树种树干液流速率与气候因子的数学拟合模型Tab.4 Mathematical fitting model of fluid flow rate and climate factor of different tree species 序号树种模型形式R21黄樟Y=-110.037-0.967 X1+0.008 X2+5.205 X3+1.134 X4+0.187 X5-0.176 X6-0.378 X70.7042马尾松Y=-73.633+3.32 X40.0383柏木Y=-2.009+0.011 X2+0.038 X3+0.041 X4+0.12 X5+0.008 X6+0.002 X70.2044杉木Y=-3.311+0.034 X2+0.051 X4+0.014 X6+0.002 X70.2305麻栎Y=12.609+6.557 X1+0.493 X2-36.267 X3-0.918 X4+1.79 X50.2736木荚红豆Y=12.887+0.126 X1+0.106 X2-1.162 X3-0.56 X4+0.603 X5-0.023 X6+0.002 X70.6147刺楸Y=-65.022+0.32 X2+30.06 X3-1.236 X4+0.813 X6+0.007 X70.4808朴树Y=1.747+0.071 X1+0.054 X2-0.383 X3-0.128 X4+0.001 X70.685 注:Y表示树干液流速率;X1表示空气湿度;X2表示空气温度;X3表示土壤湿度;X4表示土壤温度;X5表示风速;X6表示大气压强;X7表示太阳总辐射。F检验临界值<0.05。
4 结论与讨论
4.1 结论
(1) 武陵石漠化区8个树种树干液流速率日变化除黄樟、马尾松、柏木和杉木7月和9月均表现为单峰曲线趋势外,其余树种的表现为单峰或者双峰曲线趋势;8个树种树干液流速率峰值7月出现在11:00—14:00,9月出现在9:00—15:00。7月各树种树干液流速率最大值排序为刺楸>木荚红豆>朴树>麻栎>黄樟>马尾松>杉木>柏木;9月各树种树干液流速率最大值排序为麻栎>木荚红豆>黄樟>马尾松>刺楸>朴树>柏木>杉木。
(2) 刺楸、朴树7月的树干液流通量整体大于9月的,其它树种在7:00—19:00的树干液流通量小于9月的。
(3) 除麻栎、木荚红豆外,其它树种平均日耗水量都是7月的大于9月的。7月不同树种的平均日耗水量排序为刺楸>木荚红豆>马尾松>麻栎>黄樟>杉木>朴树>柏木;9月不同树种的平均日耗水量排序为麻栎>木荚红豆>马尾松>刺楸>黄樟>朴树>柏木>杉木。9月的平均日耗水量相对于7月的减少量最大的是杉木,达到258.16%;最小的是黄樟,为5.45%。
(4) 影响树木蒸腾速率的主导气候因子是太阳总辐射,其它多种环境因素共同影响。
4.2 讨论
水分是影响石漠化地区植被恢复和造林存活的关键因子。因此,选择合适的造林树种尤为重要。植物耗水是研究植物适应性和水分使用效率的重要指标。
据桑植县国家基本气象站记录,在监测期间,7月平均降雨量为756 mm,平均气温为28.3 ℃,平均相对湿度为76%,平均大气压强为9 687 hPa;9月平均降雨量为1 747 mm,平均气温为23.2 ℃,平均相对湿度为84%,平均大气压强为9 690 hPa。可以看出,7月的平均降雨量、平均相对湿度及平均大气压强均低于9月的,平均气温高于9月的。
多项研究[20-23]得出,树干液流速率的波动曲线为单峰或者双峰曲线,而本试验也证明,在相同气候条件下测定石漠化区8种树种的树干液流,其启动基本在7:00—9:00,8种树树干液流速率变化大部分是早晚低,日出后逐渐升高后再降低,呈单峰或双峰曲线。王立景等[24]、赵仲辉等[25]的研究得出:马尾松、杉木的树干液流速率日变化均呈单峰曲线,与本试验的研究结果一致,但马尾松的树干液流速率峰值出现时间为13:00—14:00,与本研究中峰值出现在12:00、13:00有所不同;杉木的树干液流速率峰值出现在12:00—13:00,与本研究中峰值出现在11:00、14:00也不同,这可能与观测期长短不同有关。刘鑫等[26]的研究结果显示,麻栎的树干液流速率日变化趋势,夏季表现为单峰曲线、秋季表现为双峰曲线、冬季表现为多峰曲线,峰值出现在11:00或13:00,与本试验中麻栎的树干液流速率日变化7月呈单峰曲线,9月呈多峰曲线,峰值出现在9:00、14:00不同。李成龙[27]的研究结果显示,朴树的日均树干液流通量,晴天为典型的单峰曲线,与本试验中选择典型晴天时朴树日均树干液流通量呈单峰和双峰曲线不同,这可能是观测目标树大小及试验期长短不同造成的。
不同树种的树干液流速率到达峰值和停止的时间有很明显的差异,树干液流速率峰值出现时间的早晚,反映了树种耗水速率的大小及耗水调控能力的强弱[16]。植物水分利用率越高,其抗干旱能力越强[28];植物的耗水速率越低,其水分利用效率相对较高。本研究中阔叶树的树干液流启动时间较早,针叶树的较晚,这是因为针叶树的抗旱能力强于阔叶树的,针叶树种叶片角质层较厚,可以遮挡一部分太阳辐射,保护气孔和表皮细胞,同时维管束较粗,气孔较小,减弱了在较强太阳辐射和较高温度条件下叶片的蒸腾作用,增强了抗旱能力[29]。
影响植物蒸腾耗水的因素除了其自身生物学特性外,还有光照、温湿度、降水、风力等。本试验研究了土壤温湿度、空气温湿度、太阳总辐射、大气压强等与树干液流速率间的相关性,发现影响树木蒸腾耗水的主导影响因子是太阳总辐射。这一结论与刘鑫等[23]的研究结果仅部分相似,可能与试验所用树种和选择季节不同有关。太阳辐射可以直接对气孔运动产生影响,同时引起相对湿度和空气温度的变化,使得多种环境因子共同作用于气孔运动,最后导致蒸腾速率的改变[21]。
本研究建立了8种树种的树干液流速率与气候因子的多元线性回归模型,可为预测树木耗水量和实现耗水量的时间尺度扩展等提供参考。但由于实验仪器和研究方法等存在的局限性,今后需结合树体枝干水容大小及根系吸收与供应水分能力的大小等在树种生态、生理及生物学等方面开展研究,进一步揭示树木的耗水特征及适应机制,建立植物生理指标与耗水之间的相关模型,为石漠化区抗旱树种快速有效选择提供依据,为植被构建和生态治理提供参考。
猜你喜欢
文萃报·周五版(2020年17期)2020-06-01
做人与处世(2020年8期)2020-04-26
科学导报(2019年19期)2019-09-23
读者(2019年10期)2019-05-08
家教世界·创新阅读(2019年4期)2019-05-04
作文与考试·高中版(2019年14期)2019-04-26
海峡姐妹(2017年12期)2018-01-31
家教世界·创新阅读(2017年11期)2017-11-27
储能科学与技术(2014年5期)2014-02-27
储能科学与技术(2014年1期)2014-02-27
