
贵州首次发现的日本松干蚧生物学特性研究
2022-01-05 10:31田茂娟喻三鹏张念念邱建生樊中亮吴晓敏王戍勃
环境昆虫学报 2021年6期
田茂娟,喻三鹏,张念念,邱建生*,樊中亮,吴晓敏,周 倩,王戍勃,李 巧
(1. 贵州省林业科学研究院,贵阳 550011;2. 贵州大学林学院,贵阳 550025;3. 汇川区林业局,贵州遵义 563000;4. 绥阳县林业局,贵州遵义 563300;5. 西南林业大学,昆明 650000)
日本松干蚧Matsucoccusmatsumurae(Kuwana.)又名黑松干蚧、赤松干蚧、松虱,属半翅目Hemiptera松干蚧科Matsucoccidae松干蚧属Matsucocus(Foldi,2004;杨钤等, 2013),是一种严重为害松属植物的外来入侵种。该蚧虫于1942年在我国辽宁旅顺被首次发现(刘卫敏等, 2015),至2017年,我国已在河北、辽宁、吉林、江苏、浙江、山东、广东、四川、陕西、安徽和上海等11省市被发现,发生面积41 203 ha(赵石峰等, 1990; 国家林业和草原局森林和草原病虫害防治总站, 2019)。目前,日本松干蚧已成为严重危害我国松属植物的重要外来有害生物(焦守武和夏鲁青, 1991; 李娟等, 2013),给我国的经济及生态带来了重大损失(万方浩等, 2015),长期被我国列为检疫性有害生物。在国外,日本松干蚧已入侵至韩国、美国、乌克兰及俄罗斯(Foldi, 2004),成为世界性入侵蚧虫。
松干蚧科Matsucoccidae现存的所有蚧虫,只在松属植物干、枝、梢的皮下及针叶上取食。日本松干蚧的主要寄主有赤松Pinusdensiflora、红松P.koraiensis、马尾松P.massoniana、华山松P.armandii、樟子松P.sylvestrisvarmongolica、油松P.tabuliformis、黑松P.thunbergii(李刚等, 2016; 国家林业和草原局森林和草原病虫害防治总站, 2019; Choietal., 2019)。我国在日本松干蚧生物学特征方面做了大量的研究,孙胜利和蒋永正(1990)研究了日本松干蚧的发生与物候现象的关系,并指出日本松干蚧的发育受温度、湿度、风雨等因素的影响。袁福香等(2014)研究表明,东北地区冬季气温上升有利于日本松干蚧的扩散、虫口密度增加。越夏期,温度为25℃时,日本松干蚧发育最快,温度为10℃时,日本松干蚧无法完成正常生长发育和繁育下一代。刘卫敏等(2015)研究了辽宁抚顺、山东青岛和浙江金华三个气候区域日本松干蚧的形态及习性,研究显示,不同气候区的各虫态发育历期明显不同,日本松干蚧在浙江金华3月上旬开始出蛰,山东青岛和辽宁抚顺的日本松干蚧于4月上旬至5月上旬开始出蛰,辽宁抚顺种群在10月份开始越冬,而浙江金华的是在12月份开始越冬。由于我国南北气候差异较大,其在不同区域的生物学习性有所不同(刘卫敏等, 2015),贵州处于北亚热带、中亚热带的交汇区域,与日本松干蚧入侵区域辽宁(中温带和暖温带)、山东(暖温带)、浙江(北亚热带和中亚热带)等地不同,因此研究其在贵州的生物学特性十分必要,以便对日本松干蚧的监测和防治提供更多参考信息。
1 材料与方法
1.1 研究地点
研究地点位于贵州省遵义市汇川区及绥阳县,汇川区最早于2014年发现马尾松不明原因死亡,发生面积为46 ha,一直误诊为真菌病害,2019年确定为日本松干蚧。所有标本及研究材料均采至遵义汇川区团泽镇(107.017°E, 27.835°N)、绥阳县蒲场镇(107.043°E, 27.872°N)日本松干蚧发生区的马尾松林分,树龄15~20 a,树高10~20 m,胸径10~20 cm。
1.2 方法
形态鉴定:通过采自松干蚧发生区的雌成虫、雄成虫与Foldi和Morrison描述的日本松干蚧的触角、背疤、胸气门、翅、爪等进行形态对比(Morrison, 1928; Foldi, 2004),确定其分类地位。
生活史观察:从2019年4月到2021年4月连续开展观察,每20~30 d伐有受害状的马尾松3株进行每木检查,观察统计各虫态虫口密度情况,拍摄生态照片;通过野外观察和室内培养方式,记录日本松干蚧从卵期到成虫到交配生殖的整个生活史行为过程并测定其不同龄期虫体数据。
孵化率统计:取完整卵囊放入培养皿,在培养皿壁上均匀涂抹凡士林,室温(15~24℃),相对湿度70%~90%,每日观察孵化情况直至卵孵化完毕,统计凡士林上若虫数及卵囊中未孵化的卵数量,计算孵化率。
2 结果与分析
2.1 在贵州发现的日本松干蚧各虫态的形态特征
将采自汇川区和绥阳县的松干蚧雌成虫(图1)、雄成虫(图2)、3龄雄若虫(图3)、初孵1龄若虫(图4)与Foldi、Morrison描述的日本松干蚧Matsucoccusmatsumurae(Kuwana, 1905)雌成虫、雄成虫(Morrison, 1928; Foldi, 2004)及其它虫态,进行形态对比研究,发现形态特征与日本松干蚧一致。结合其生活史为一年2代,雌虫未经交配不能产卵,没有孤雌生殖现象,将其与马尾松干蚧Matsucoccusmassonianae、云南松干蚧Matsucoccusyunnanensis区分,确定为日本松干蚧M.matsumurae(Kuwan),每个虫态的形态特征描述如下。
雌成虫:卵圆形,长2.20±0.225 mm (n=17),宽1.24±0.266 mm (n=17),橙红或橙褐色,体璧柔韧,体节不明显,头端较窄,后部肥大,腹部末端有倒“V”型臀裂。单眼黑色,1对。胸足转节三角形,转节上有1根长刚毛;腿节粗,胫节略弯;跗节1节,基部1/4处有缢缩,外观看似2节,端部有爪,爪基部有端部膨大成冠状的爪跗毛1对,长度超过爪端。触角9节,长0.72±0.075 mm (n=24),基部两节(柄节、梗节)粗大,其余各节(鞭节)为念珠状,6~9节各有2根粗壮感觉器。背疤成排分布,数量在200~300个,第Ⅷ腹节也有少量背疤。多格腺集中在最后一节,成一群,数量40~60个,多格腺内双孔,外部有12个小孔(图1)。

图1 日本松干蚧雌成虫形态特征(王戍勃和喻三鹏摄)Fig.1 Morphological characteristics of female adults of Matsucoccus matsumurae (Photoed by WANG Shu-Bo and YU San-Peng)注:A,雌成虫;B,转节;C,刚毛;D,背疤;E,多格腺结构;F,多格腺;G,6~9节2根感觉器;H,触角;I,单眼、倒“V”型臀裂;J,足;K,爪及爪跗毛;L,跗节。Note: A, Female adults; B, Trochanter; C, Setae; D, Back scar; E, Multigrid glandular structure; F, Multigrid glands; G, 6~9 segments with 2 sensory organs; H, Antenna; I, Monocular, inverted "V" hip split; J, Legs; K, Claws and tarsal hairs; L, Tarsal joints.

图2 日本松干蚧雄成虫形态特征(喻三鹏摄)Fig.2 Morphological characteristics of male adults of Matsucoccus matsumurae (Photoed by YU San-Peng)注:A,雄成虫;B,翅脉;C,翅羽状纹;D,勾状交尾器;E,马蹄片及柱状管腺;F,足;G,头、复眼及触角;H,触角;I,腹部、马蹄片及长蜡丝。Note: A, Male adult; B, Wing vein; C, Wing pinnate pattern; D, Hook-shaped copulatory apparatus; E, Horseshoe piece and columnar tube gland; F, Foot; G, Head, compound eye and antennae; H, Antennae; I, Abdomen, horseshoe piece and long wax silk.
雄成虫:长1.18±0.96 mm (n=10),宽0.39±0.023 mm (n=10)。头、胸部黑褐色,腹部淡褐色。触角丝状,长1.25±0.088 mm (n=20),10节,基部两节粗短,其余各节细长,生有许多刚毛,复眼大而突出,紫褐色,口器退化,胸部膨大,足细长。前翅发达,长1.61±0.084 mm(n=18),宽0.68±0.079 mm (n=18),膜质半透明,翅面有明显的羽状纹;后翅退化为平衡棒,端部有丝状钩刺3~7根。腹部9节,第7节背面有1个马蹄形的硬片,其上柱状管腺10~18根,分泌白色长蜡丝,长度为2.23±0.258 mm (n=10)。腹部末端有1个钩状交尾器,向腹面弯曲(图2)。
3龄雄若虫的外形与雌成虫相似,但其个体小于雌成虫,长1.61±0.247 mm (n=24),宽0.91±0.246 mm (n=24),椭圆或卵圆形,红褐色,触角9节,长0.55±0.111 mm (n=48),基部2节粗大,其余各节为念珠状。腹部末端无倒“V”型臀裂。具爪及爪跗毛1对,长度超过爪端(图3)。

图3 日本松干蚧3龄雄若虫的形态特征(喻三鹏摄)Fig.3 Morphological characteristics of the 3rd instar male nymphof Matsucoccus matsumurae (Photoed by YU San-Peng)注:A,外观;B,触角;C,爪及爪跗毛。Note: A, Appearance; B, Antennae; C, Claws and hocks.

图4 日本松干蚧初孵若虫的形态特征(喻三鹏摄)Fig.4 Morphological characteristics of newly hatched nymphsof Matsucoccus matsumurae (Photoed by YU San-Peng) 注:A,外观;B,触角;C,尾丝。Note: A, Appearance; B, Antennae; C, Tail.
初孵若虫:从形态特征上很难将雌雄区分,梭形,淡黄色,长0.33±0.019 mm (n=21),宽0.13±0.01 mm (n=21)。触角6节,长0.09±0.006 mm (n=41),柄节宽大有1根短刚毛,第2节有2根短刚毛和1根长刚毛,第三节较小,念珠状,两边各有1宽感觉器,第4节有1弯曲的肉质状刚毛,第5节较小念珠状无刚毛,第6节顶端有一指形突,看似7节,有4根刚毛,1对肉质感觉器。腹部末端有刚毛2对,一长一短,长刚毛长度0.16±0.006 mm (n=41)。足细长,跗节1节,端部有爪,爪基部具冠状爪跗毛1对。
2.2 日本松干蚧在贵州区域的生活周期
日本松干蚧为渐变态昆虫,其中雌虫为渐变态,发育过程包括若虫(2龄)和成虫两个阶段;雄虫为过渐变态,发育过程经历若虫(3龄)、蛹和成虫3个阶段(图5)。

图5 日本松干蚧生活周期Fig.5 Life cycle of Matsucoccus matsumurae (Kuwana.)
该虫以有性繁殖繁育后代。在贵州1年发生2代,第一代为3月中下旬至10月中下旬;第二代为9月中旬至次年5月中下旬,以第二代1龄寄生若虫越冬(表1)。越冬的1龄寄生若虫在3月上中旬出蛰,蜕皮后进入2龄,附肢消失,成为珠形无肢若虫,后体型逐渐膨大,初期为扁球形,后成圆球形,并逐渐从树皮缝中显露出来,进入展露期。这一阶段,在干、枝上可明显看到圆球形或椭圆球形2龄若虫珠形体,颜色为黄褐色至棕褐色,边缘有排列整齐的蜡丝。2龄雌雄若虫在个体上差异明显,雌若虫个体比雄若虫大。3月中旬,2龄雄若虫经过一段时间发育后,开始从珠形体中陆续破壳而出,成为能够爬行的3龄雄若虫,其整体呈现黄褐色,触角和足为黄色,整个身体从壳中出来后,便迅速爬行,寻找干、枝隐蔽处固定,从身体各部分泌蜡丝,将身体包裹成茧,身体萎缩然后开始化蛹,茧长1.74±0.201 mm (n=10),宽0.83±0.135 mm (n=10)(图6)。2龄雌若虫珠形体继续发育。3月中下旬,部分雌成虫开始出壳,少数雄成虫也开始羽化,雌雄成虫开始交配,雌成虫开始产卵。分钟不等。雄成虫交配后几小时即死亡,未经交配的雄成虫可以存活2~5 d,1头雄成虫最多可以与2头雌成虫交配。雌成虫交配后,15~30 min便能够爬到树干、枝的翘裂皮下,粗老皮缝,枝的基部及球果鳞片等隐蔽处分泌蜡丝形成卵囊,雌成虫在卵囊中可存活2~3 d。每头雌成虫平均产卵228±38粒(n=12)。卵囊椭圆形,长0.20±0.014 mm (n=20),宽0.13±0.004 mm (n=20),包被较为疏松,能够随风飘扬。卵产于雌成虫分泌蜡丝形成的白色絮状卵囊中,椭圆形,浅黄色到橙黄色(图7、图8)。

表1 日本松干蚧在贵州的生活史Table 1 Life history of Matsucoccus matsumurae (Kuwana.) in Guizhou

图6 日本松干蚧雌雄若虫形态(邱建生摄)Fig.6 Morphology of male and female nymphs of Matsucoccus matsumurae(Photoed by QIU Jian-Sheng)注:A,2龄雌雄若虫;B,3龄雄若虫;C,3龄雄若虫及结茧。Note: A, 2nd male and female nymphs; B, 3rd male nymphs; C, 3rd male nymphs and cocoons.

图7 日本松干蚧雌成虫脱壳、交配及产卵过程(邱建生和喻三鹏摄)Fig.7 Shelling, mating and oviposition process of female adults of Matsucoccus matsumurae (Photoed by QIU Jian-Sheng and YU San-Peng)注:A,雌成虫出壳;B,雌成虫爬行;C,雌雄交配;D,交配后雌成虫分泌蜡丝结卵囊。Note: A, Female adults came out of shell; B, Female adults crawled; C, Male and female mated; D, Female adults secreted wax silk knot oocysts after mating.

图8 日本松干蚧卵囊及卵形态(邱建生摄)Fig.8 Oocystsand egg morphology of Matsucoccus matsumurae (Photoed by QIU Jian-Sheng)注:A,雌成虫;B,卵囊;C,卵。Note: A, Female adults; B, Oocysts; C, Eggs.
4月中下旬,卵孵化,卵囊内的若虫2~3 d可孵化完毕,孵化率可达97.90%,第一代若虫产生。初孵若虫能快速爬行,迅速扩散,1~2 d后,便转入树干、枝的翘裂皮下和缝隙中,开始固定取食,体形逐渐转变为珠形,虫体周边逐渐分泌出白色蜡丝,呈放射状成对排列在身体两侧,种群进入寄生若虫阶段。此阶段,种群整体进入隐蔽期,虫体较小,均在树干、枝的皮下,很难发现。从5-8月上旬,1龄寄生若虫均处于树皮下隐蔽越夏。
8月中下旬,处于隐蔽期的1龄若虫开始脱皮进入2龄,体型逐渐增大,从树皮下逐步显现,种群进入显露期。经过一段时间发育后,3龄的雄若虫开始从2龄珠形体破囊而出,爬行寻找作茧化蛹场所,很快固定,从身体各部分泌蜡丝,将身体包裹起来成茧,自身萎缩成预蛹然后成蛹,7~10 d后羽化,寻找雌成虫交配,后死亡,雄性成虫寿命最短为5 h,最长为2 d(图9、图10)。

图9 日本松干蚧雄虫各时期虫态(邱建生摄)Fig.9 Males at different stages of Matsucoccus matsumurae(Photoed by QIU Jian-Sheng)注:A、B,茧、蛹;C、D,羽化、雄成虫。Note: A, B, Cocoon, pupa; C, D, Eclosion, male adult.

图10 日本松干蚧雄成虫形态(邱建生摄)Fig.10 Male adult morphology of Matsucoccus matsumurae (Photoed by QIU Jian-Sheng)注:A,B,C,雄成虫。Note: A, B, C, Male adults.
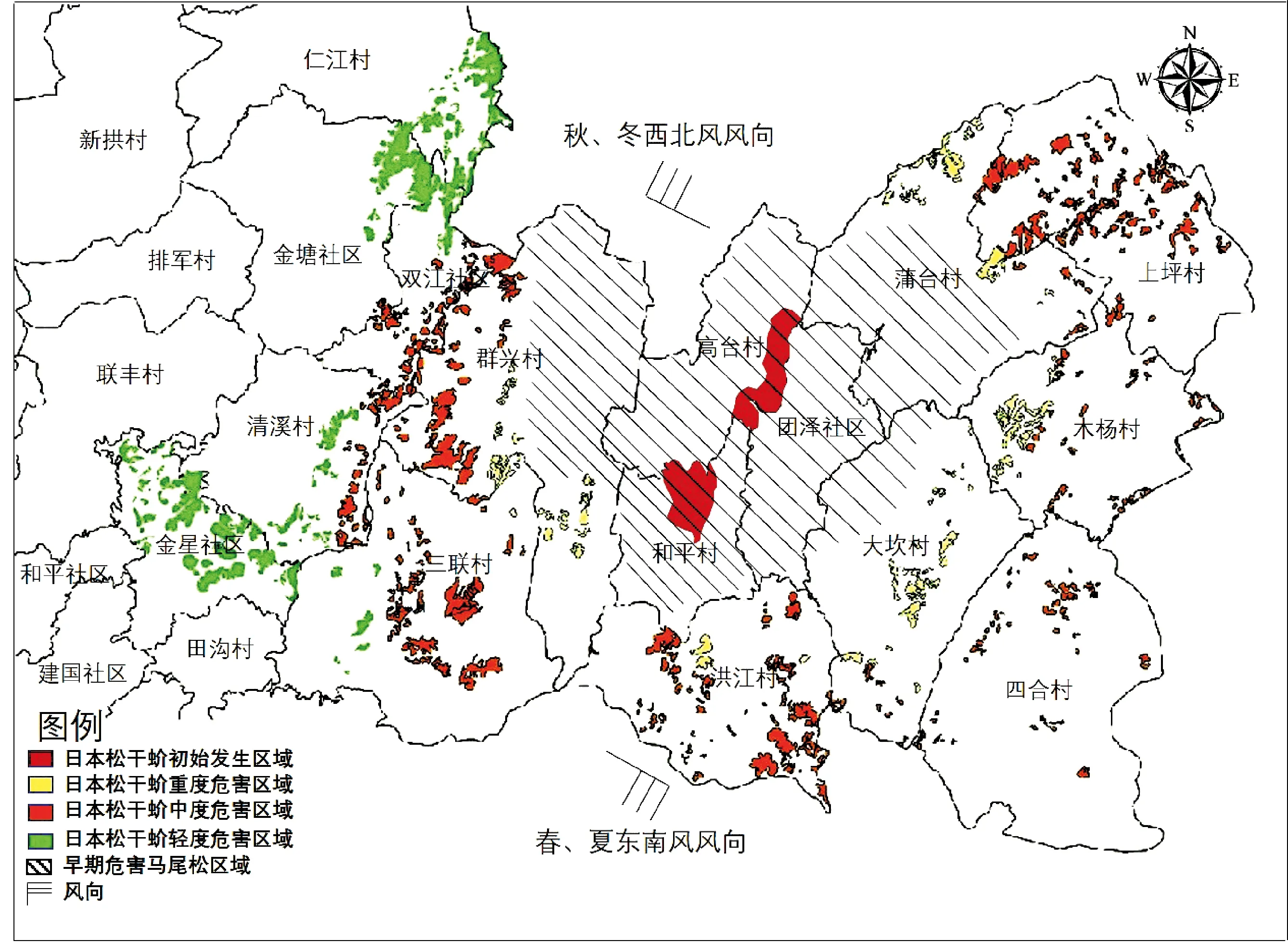
图11 日本松干蚧在汇川区的传播图Fig.11 Spread of Matsucoccus matsumurae in Huichuan District
雄虫羽化同时,雌成虫也从2龄珠形体中破壳而出,四处爬行,寻找雄成虫交配,然后寻找树干、枝及梢等隐蔽处固定,很快分泌蜡丝包裹身体形成卵囊。雌成虫平均产卵228±38粒(n=12)。卵在9月上中旬开始孵化,9月下旬至10上旬达到高峰。第二代初孵若虫活动快速,四处扩散寻找树干、枝的皮下固定取食,形成梨形无肢若虫,隐蔽越冬,直到次年3月中上旬出蛰,开始新的循环。
2.3 日本松干蚧在贵州区域的危害与传播情况
从2014年在汇川区发现松干蚧危害以来,由于前期误诊为真菌病害,导致日本松干蚧在贵州遵义市汇川区、绥阳县及新蒲新区不断扩散蔓延,到2020年发生面积已达2 096 ha,中度以上发生面积1 883 ha,占89.84%。已有270 ha松林死亡,占12.91%,共导致62 993株马尾松枯死。日本松干蚧是首次在贵州发现,也是首次在西南区域发现,表明该虫完全能够在西南地区建立种群并迅速传播蔓延,也意味着日本松干蚧已进入到北纬27°范围。
目前,日本松干蚧在贵州遵义仅发现危害马尾松,而附近的湿地松Pinuselliottii、华山松未见危害。春、秋两季的风是其在林分间和植株间做短距离自然传播的主要动力,雨水亦能把卵囊、若虫及成虫冲刷至低洼处,还可附于昆虫和鸟类体上带至其它地方(蔺成阁等, 2011; 徐云峰等, 2014; 任卫红和赵恒刚, 2018)。从汇川区日本松干蚧受害马尾松林分分布图(图11)可以看出,日本松干蚧早期危害区域大部分马尾松已死亡,目前受害马尾松林分主要分布在最初发源地高台村、和平村的两侧,说明日本松干蚧有两个传播方向,西北向和东南向,春季日本松干蚧第一代卵及初孵若虫受东南风吹拂向西北方向传播;秋季第二代卵及初孵若虫受西北风吹拂向东南方向传播,进而形成目前的分布态势。日本松干蚧在汇川区发生的10年间,向西传播了14 km;向东传播了10 km,传播面积已达2 096 ha。
卵囊被风带到其它马尾松上后,若虫从其中孵出并开始隐蔽危害马尾松,随着种群不断繁殖,松树开始出现部分枯死枝,随后枯死枝由下而上不断增多,直至最后整株枯死。通常从松树出现枯死枝到整株枯死,时间为1年。由于日本松干蚧多集中在枝及枝梢的阴面危害,常造成受害马尾松出现主梢和枝弯曲现象,可以用作判断松干蚧危害的依据(图12)。

图12 马尾松受害状Fig.12 Injury of Pinus massoniana Lamb.注:A,马尾松单株受害状(初期);B,马尾松群体受害状(中期);C,马尾松顶梢弯曲。Note: A, P.massoniana single plant injured (early); B, P.massoniana population injury (metaphase); C, P.massoniana tip bending.
3 结论与讨论
对贵州省遵义市汇川区和绥阳县马尾松林分发生的松干蚧雌雄成虫进行分类鉴定,结果表明,其主要形态特征均符合日本松干蚧的分类特征,结合其生物学特性,确定为日本松干蚧M.matsumurae(Kuwana.)。日本松干蚧在贵州一年发生2代,第一代为3月中下旬至10月中下旬,第2代为9月中下旬至次年5月中下旬,以第2代1龄寄生若虫越冬。以有性生殖繁育后代,雌成虫平均产卵年为228±38 (n=12)粒,孵化率达97.90%;未见孤雌生殖现象。日本松干蚧目前在贵州遵义市汇川区、绥阳县、新蒲新区不断扩散蔓延,但仅发现危害马尾松,到2020年,危害面积已达2 096 ha,共导致62 993株马尾松枯死。日本松干蚧主要依靠风力做短距离自然传播,春季主要依靠东南风往西北方向传播,秋季主要依靠西北风往东南方向传播。
日本松干蚧在贵州区域一年发生2代,与浙江、山东、辽宁等地发生的世代数一致,但其越冬和越夏时间有所不同,发生在贵州的日本松干蚧于5月进入越夏,11月进入越冬阶段,辽宁抚顺地区于10月份、浙江金华地区于12月份进入越冬阶段,这符合不同地区世代发生时间略有差异的特点(蔺成阁等, 2011; 刘卫敏等, 2015; 孟庆兰等, 2016)。此次在贵州区域发现日本松干蚧,表明其已进入西南区域,并跨越到北纬27°范围,整体向南又推进了3个纬度,我们注意到2014-2017年中国林业有害生物普查成果中,四川巴中市和通江县有日本松干蚧分布记录(国家林业和草原局森林和草原病虫害防治总站, 2019),其刚好与贵州遵义发生的日本松干蚧处于同一经度范围内,说明日本松干蚧在西南地区传播范围广、尺度大。贵州的黔中和黔北区域为马尾松的集中分布地,日本松干蚧的传入黔北后将对这一区域的马尾松林分构成重大威胁。
贵州发现的日本松干蚧在外形上要小于其他区域的日本松干蚧,杨平澜等(杨平澜等, 1976)的描述中,初孵若虫体长0.362~0.400 mm,宽0.150~0.175 mm,雌成虫体长约4 mm,雄成虫体长约2 mm,在本研究中,初孵若虫体长0.33±0.019 mm,宽0.13±0.01 mm,雌成虫体长2.20±0.225 mm,雄成虫体长1.18±0.96 mm,这可能是入侵环境对其影响的原因,万方浩等(2015)研究表明入侵种群的可塑性程度更高。在贵州遵义入侵的日本松干蚧体长在2.0~3.1mm之间,在形态上更接近于英国松干蚧Matsucoccuspini(Green),单从形态上很难确定其归属,最终根据其第8腹节分布有背疤、寄主情况及生活史,才将其最终确定为日本松干蚧M.matsumurae(Kuwana.)。研究发现日本松干蚧的雌成虫不交配是不能产卵的,说明其以有性生殖繁育后代,没有发现孤雌生殖现象,这也与云南松干蚧M.yunnanensisFerris有所不同,由此,可以看到,日本松干蚧的雌雄分化在卵期即以确定。为此,本研究在文中将其生活史图进行了重新描绘,使其更能反映该虫在自然环境的生活本质。但1龄若虫在外形上很难区分雌雄,且在松干蚧属内的不同种间,1龄若虫在形态上非常相似,本研究对比了云南松干蚧M.yunnanensisFerris和法国松干蚧M.feytaudiDucasse的1龄若虫(Foldi, 2004),发现其无论在外观形态、触角节数、每节的大小及刚毛数、尾部刚毛等基本都是相同的,这些现象都是造成我国早期松干蚧分类混乱和争论不断的客观原因,因此,国内在松干蚧的形态分类仍然需要建立更完善的体系,逐渐量化各分类特征,进行更多的比较解剖学的研究,特别是在分子水平上开展更多的工作,这都是下一步急需努力的方向。
猜你喜欢
农业工程学报(2022年10期)2022-08-22
天津农业科学(2022年5期)2022-05-31
少先队活动(2021年5期)2021-07-22
饲料博览(2021年1期)2021-04-05
饲料博览(2020年11期)2020-12-30
新农民(2020年15期)2020-06-22
晚晴(2020年12期)2020-05-18
中国科技纵横(2017年13期)2017-08-09
热带农业工程(2016年6期)2017-05-17
晚晴(2016年6期)2016-05-14
