
RNA结合蛋白CELF4对小鼠早期胚胎发育的作用
2022-01-18 06:50史河秀钟玉环刘玥孙建东莫开恩王世鄂
中国组织化学与细胞化学杂志 2021年4期
史河秀,钟玉环,刘玥,孙建东,莫开恩,王世鄂*
(1福建卫生职业技术学院医学基础部 福州 350101;2福建医科大学干细胞工程与再生医学福建省高校重点实验室 福州 350108)
现代辅助生殖技术给生育障碍夫妇带来了福音,但哺乳动物早期胚胎体外发育存在发育阻滞现象,导致人工体外受精成功率不高,因此深入了解胚胎发育机制至关重要。RNA结合蛋白可与多种靶RNA结合,参与RNA成熟、转运、定位和翻译等过程。研究表明,RNA结合蛋白(RNA-binding proteins,RBPS)在胚胎发育中具有重要作用[1]。CUG-BP and Elav 样 家 族(CUG-BP and Elav-like family,CELF)是重要的RNA结合蛋白,具有6个成员(CELF1-6),因与果蝇Bruno蛋白具有很高的同源性,也被称作Bruno-like (Brunol)家族。CELF可参与前体mRNA的选择性剪接、胞嘧啶到尿嘧啶的RNA编辑(C-U-RNA编辑)和调节mRNA稳定性和翻译等过程。已证实CELF在配子发生和早期胚胎发育、心脏和骨骼肌功能以及神经突触传递等方面具有重要作用[2,3],如:Celf1敲除小鼠可导致配子发生缺陷[4]。对CELF成员CELF4的研究目前主要集中在神经系统方面:CELF4基因突变可导致癫痫发作[5],其单倍体不足与儿童自闭症相关[6]。我们先前研究发现,CELF4分布于小鼠卵巢的卵母细胞中,提示CELF4可能与早期胚胎密切发育有关[7]。但CELF4在早期胚胎发育中的作用尚未见报道。
材料与方法
1 实验动物和主要试剂
SPF级昆明(Kunming,KM)小鼠(雌鼠4~6周,雄鼠8周以上)。孕马血清促性腺激素(PMSG)(宁波激素制药二厂),人绒毛膜促性腺激素(hCG)(ProSpec TechnoGene),M2培养液(Sigma),KSOM培养液(Millipore),聚乙烯醇粉末(PVA粉 末,Sigma)、台 氏 液(Tyrode’s solution,Sigma),兔抗CELF4(BRUNOL4)多克隆抗体(Santa Cruz),Alexa Fluor® 594标记驴抗兔二抗(Life Technology),兔IgG(北京碧云天),DAPI(北京碧云天),Quick-RNA™ MicroPrep R1050试剂盒(Zymo Research),逆转录试剂盒(Thermo,SYBR® Premix Ex TaqTM(Roche)。
2 鼠早期胚胎收集
KM雌鼠腹腔注射 10 IU PMSG,48h后注射 6 IU hCG,随后均与KM雄鼠 1:1合笼,次晨检查阴栓,有阴栓者供本实验使用。分别于hCG注射后27h、42h、54h、64h、72h、90h,用M2培养液从输卵管(前期阶段)或子宫(桑椹胚和囊胚)中冲洗获得早期胚胎。
3 激光扫描共聚焦显微镜检测
上述收集到的各阶段早胚,于0.1% PVA-PBS清洗3次,台式液中去除透明带,室温固定1h,0.5%Triton X-100通透30min,0.2% BSA封闭1h,兔抗CELF4多克隆抗体(1:800稀释)4℃孵育过夜,Alexa Fluor® 594标记驴抗兔二抗(1:1000稀释)避光孵育1h,DAPI(1μg/ml)避光孵育1h,激光扫描共聚焦显微镜观察。
4 微注射操作和早胚培养
hCG注射后18h收集1-细胞胚,在KSOM中培养3h后,挑取有明显雌、雄原核的1-细胞胚,分为KSOM培养组(空白对照组)、兔IgG(100μg/ml)注射组(阴性对照组)和CELF4抗体(100μg/ml)注射组。用显微注射仪分别往1-细胞胚注射100μg/ml CELF4抗体和100μg/ml兔IgG,注射后移入KSOM培养液,在37℃、5%CO2饱和湿度的培养箱中连续培养,在hCG注射后43h、66h、96h和118h分别观察2-细胞胚、4-细胞胚、桑椹胚和囊胚发育率。
5 实时定量PCR
上述收集到的各阶段早胚或显微注射操作过的早胚,每组50枚按Quick-RNA™ MicroPrep R1050试剂盒步骤提取RNA,按Thermo K1622 逆转录试剂盒合成cDNA,按SYBR® Premix Ex TaqTM试剂盒Real-Time PCR反应。
6 计学处理
实时定量PCR数据利用2-ΔΔCt法处理mRNA数据,利用GraphPAD Prism8.0作图,利用SPSS18.0对实验数据做方差齐性检验后进行独立样本t检验;显微注射操作后细胞培养数据利用SPSS18.0软件进行卡方检验;以P<0.05为差异具有统计学意义。
结 果
1 CELF4在小鼠早期胚胎中的分布
激光扫描共聚焦显微镜观察显示:CELF4蛋白呈现红色荧光,细胞核为蓝色荧光,CELF4呈颗粒状弥散分布于各阶段早期胚胎的细胞质内,且在8-细胞胚和桑椹胚中,颗粒变粗荧光变强,但在囊胚阶段荧光则明显减弱(图1)。

图1 CELF4在小鼠早期胚胎的分布。比例尺,50μmFig. 1 The distribution of CELF4 in mouse early embryos. Scale bar, 50μm
2 CELF4 mRNA在小鼠植入前胚的表达水平
实时定量PCR检测显示,CELF4 mRNA在1-细胞胚、2-细胞胚和4-细胞胚表达较高,从8-细胞胚至囊胚阶段则表达显著下降,囊胚阶段几乎检测不到CELF4 mRNA(图2)。
3 CELF4抗体抑制囊胚发育率
往1-细胞胚注射100μg/ml CELF4抗体和100μg/ml兔IgG后,移入KSOM培养液,在hCG注射后43h、66h、96h和118h分别观察各注射组和KSOM空白对照组的2-细胞胚、4-细胞胚、桑椹胚和囊胚发育率。结果显示,KSOM组和IgG组各期早胚发育率无显著差别,CELF4抗体注射组与KSOM组和IgG组的2-细胞胚、4-细胞胚和桑椹胚的发育率没有显著变化,但CELF4抗体注射组的囊胚发育率比KSOM组、IgG组显著降低(图3,表1)。
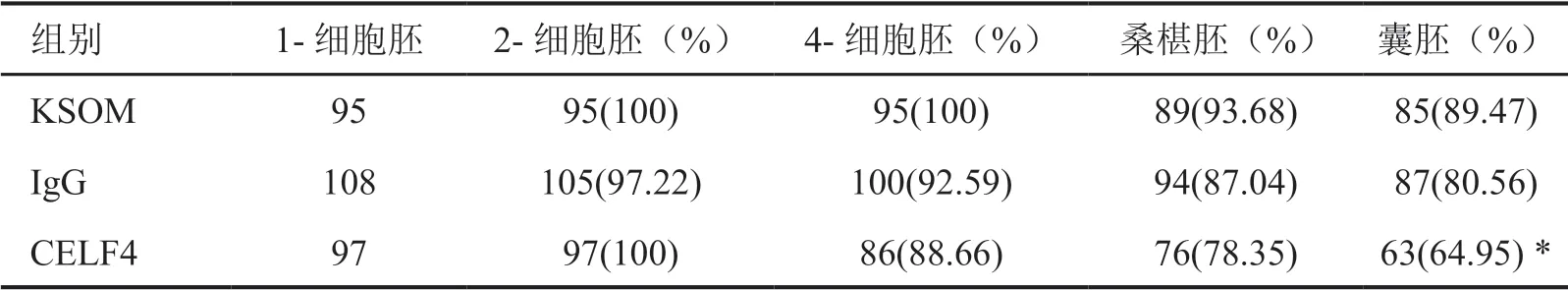
表1 CELF4抗体封闭后对小鼠早期胚胎发育的影响Tab. 1 The effect of CELF4 antibody on the development of mouse early embryos
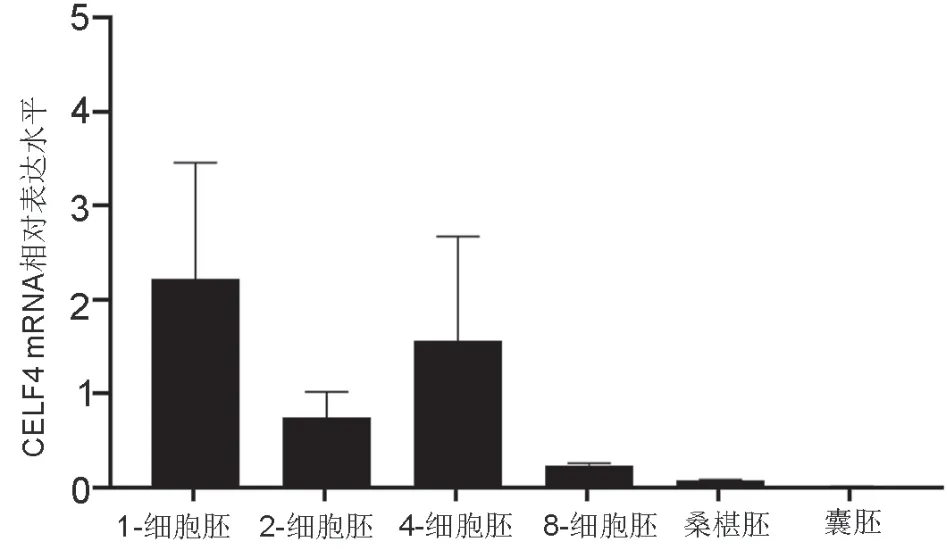
图2在小鼠早期胚胎CELF4 mRNA表达Fig. 2 CELF4 mRNA expression in mouse early embryos

图3 CELF4抗体封闭后对小鼠早期胚胎发育的影响。比例尺,50μmFig. 3 The effect of CELF4 antibody on the development of mouse early embryos. Scale bar, 50μm
4 CELF4抗体封闭后对2-细胞胚MuERV-L、eIF1A、Hsp70.1和Zscan4d mRNA的表达无影响
对经显微注射体外培养至hCG后43h的2-细胞胚进行实时定量PCR检测显示:与IgG组和KSOM组对比,CELF4抗体注射组的MuERV-L、eIF1A、Hsp70.1和Zscan4d mRNA表达变化均无统计学意义(见图4)。

图4 体外注射CELF4抗体对2-细胞胚期MuERV-L、eIF1A、Hsp70.1和Zscan4d mRNA的影响Fig. 4 Effect of CELF4 antibody injection on the mRNA expression of zygotic gene activation marker genes (MuERV-L, eIF1A, Hsp70.1 and Zscan4d ) in mouse 2-cell embryos in vitro
5 CELF4抗体抑制桑椹胚干细胞Cdx2 mRNA表达
对经显微注射体外培养至hCG后96h的桑椹胚进行实时定量PCR检测显示:与IgG组和KSOM组对比,CELF4抗体注射组Oct4、Sox2 mRNA表达水平并无明显差异,而Cdx2 mRNA表达则显著下调(图5)。
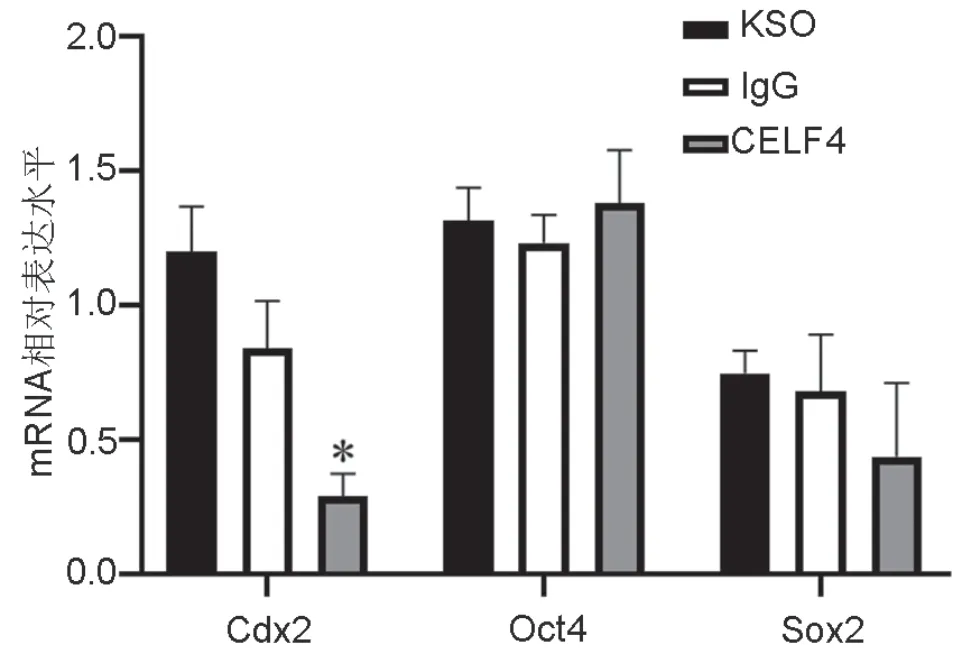
图5 CELF4抗体对桑椹胚中Cdx2、Oct4和Sox2 mRNA表达水平的影响。 *,与IgG组和KSOM组相比,P < 0.05Fig. 5 Effect of CELF4 antibody injection on the mRNA expression of Cdx2, Oct4 and Sox2 in the morula. *, P < 0.05, compared with IgG group and KSOM group
讨 论
研究表明,细胞中RNA可与RNA结合蛋白(RBPs)结合形成核糖核蛋白(RNP)复合物,RBPs通过影响RNA的结构,在生物发生、稳定性、功能、运输和细胞定位方面发挥关键作用。CELF作为RNA结合蛋白,在细胞质中可控制靶RNA的翻译或者稳定性。本研究发现CELF4颗粒状弥散分布于小鼠各阶段早期胚胎的细胞质内,且8-细胞胚和桑椹胚中颗粒变粗荧光变强,提示CELF4可能参与调控小鼠囊胚形成,其调控机制可能与CELF4将靶mRNA隔离到颗粒中调控其表达相关。
本研究观察到:CELF4 mRNA在1-细胞胚、2-细胞胚和4-细胞胚表达较高,8-细胞胚至囊胚阶段显著下降,囊胚阶段几乎检测不到。CELF4 mRNA的表达规律与CELF4蛋白的表达类似,但mRNA表达下降时间较蛋白表达时间早,这说明Celf4基因表达存在“转录-翻译非同步”现象[8]。CELF4抗体显微注射小鼠1-细胞胚后,囊胚发育率显著降低,进一步证明CELF4与囊胚发育密切相关。
哺乳动物早期胚胎发育主要经历包括合子基因组激活、致密化以及囊胚腔化等重要生理事件[9]。卵母细胞在发育过程中积累了大量母源的RNA和蛋白质支持受精后的胚胎继续发育,随着发育进行,母源物质逐渐被消耗降解,合子基因组激活(zygotic gene activation, ZGA),转为由合子型的RNA和蛋白质来支持胚胎继续发育。小鼠大规模ZGA发生在2-细胞胚期,若ZGA失败,则发生“2-细胞阻滞”。MuERV-L基因在1-细胞胚S期即产生转录活动,在2-细胞胚呈现瞬时高表达[10]。eIF-1A在小鼠2-细胞胚出现短暂高水平的转录活动[11]。Hsp70.1在小鼠ZGA早期即大规模转录[12]。Zscan4d在小鼠早期胚胎中只在2-细胞胚晚期阶段发生转录活动[13]。MuERV-L, eIF1A, Hsp70.1, Zscan4d表达于ZGA期间,它们的转录受到精确调节,因此作为ZGA的标志基因。若它们在ZGA期转录水平异常,可视为ZGA失败。本研究检测上述4个基因在CELF4抗体注射后的表达水平变化,结果发现其表达无显著差异,提示CELF4可能不通过影响合子基因组激活过程参与调控囊胚形成。
桑椹胚和向囊胚的顺利过渡是早期胚胎发育中的另一重要事件。Cdx2、Oct4和Sox2是小鼠内细胞团和滋养外胚层分化的重要调节因子[14],能促进胚胎干细胞的自我更新和多能性,并维持基因组的完整性。Oct4是POU转录因子家族的一员,在体外,转录因子Oct4是胚胎干细胞的建立和维持所必需的,在体内,它阻止早期胚胎向滋养层细胞的异位分化。通过建立母源和合子型Oct4敲除转基因小鼠模型证明,Oct4对于合子的形成、早期卵裂是必不可少的。在桑椹胚向囊胚转化过程中诱导Oct4缺失,会破坏内细胞团的谱系特异性识别能力和获得外胚层或原始内胚层的分子结构特征[15]。Sox2、Nanog和Cdx2是Oct4的靶点,三者形成网络来共同调控其他靶基因。在囊胚形成之前,HIPPO通道将SOX2限定在内细胞团干细胞中,Sox2最初通过FGF4非细胞自主地促进原始内胚层的发育,并在内细胞团中维持多能性基因的表达[16]。Cdx2是滋养层细胞分化所需的特异性转录因子,在滋养层细胞自我更新中是不可或缺的。单独敲低牛胚胎中的Cdx2或Oct4基因,另一种基因的mRNA表达受到抑制[17]。在早期胚胎中,Cdx2最初与Oct3 /4共表达,它们在胚胎干细胞中形成复合体,抑制彼此的表达[18]。将OCT4或CDX2特异性siRNAs注入牛受精卵,OCT4下调的囊胚发育率较低,CDX2和FGF4的表达也降低。CDX2下调导致囊胚腔形成延迟[19]。
本研究显示桑椹胚干细胞转录基因Oct4、Sox2 mRNA的表达无显著变化,Cdx2 mRNA的表达则显著下调。提示CELF4可能通过调控滋养层Cdx2 RNA的翻译或者稳定性从而影响囊胚的形成。Pereira等研究发现,RNA结合蛋白MEX3A可通过与Cdx2 mRNA的3’非翻译区结合,影响其表达[20]。CELF4调控Cdx2的具体机制是否与MEX3A相似,有待后续进一步研究。
猜你喜欢
鸭绿江(2021年17期)2021-10-13
鸭绿江·华夏诗歌(2021年6期)2021-09-13
中国生殖健康(2020年5期)2021-01-18
生殖医学杂志(2020年12期)2020-12-22
浙江医学(2020年17期)2020-09-21
中国生殖健康(2018年5期)2018-11-06
中成药(2017年5期)2017-06-13
新疆医科大学学报(2015年10期)2015-12-26
中国医学科学院学报(2015年5期)2015-03-01
郑州大学学报(医学版)(2015年2期)2015-02-27
