
不同树龄同心圆枣营养器官解剖结构与其抗旱性的关系*
2022-02-18 08:39赵宏亮李永华黄占明倪细炉
南方农机 2022年4期
程 昊 , 赵宏亮 , 李永华 , 黄占明 , 倪细炉
(1.宁夏大学农学院,宁夏 银川 750021;2.宁夏林业研究院种苗生物工程国家重点实验室,宁夏 银川 750004;3.宁夏大学西北土地退化与生态恢复国家重点实验室培育基地,宁夏 银川 750021;4.宁夏贺兰山森林生态系统国家定位观测研究站,宁夏 银川 750021)
枣树(Zizyphus jujubeMill.)属鼠李科枣属乔木,素有“铁杆庄稼”之称,是我国栽培历史较长的落叶果树之一[1]。其果实含丰富的蛋白质、氨基酸、多糖等养分,有较高的药用、保健作用和经济价值,受到人们的重视。同心圆枣(Zizyphus jujube cv.“Tongxinyuanzao”)原产于宁夏同心县王团镇,具有悠久的栽培历史,同时具有很强的抗逆性和适应性能力,在降雨量不足100 mm的干旱带地区能存活,是宁夏特有的极耐干旱的枣树品种,也是我国干旱、半干旱地区的经济支柱产业[2]。植物形态结构和功能的统一是植物适应干旱环境的生物学基础[3]。在形态结构方面,凡是抗旱性强的植物,植物的表面积不发达、叶面积小、表皮角质层发达、叶组织较紧密,栅栏组织和叶脉都很发达,气孔下陷、单位面积气孔数较多,保水能力强、根系发达等。长期以来,对植物旱生结构的研究主要为新疆罗布麻[4]、骆驼蓬属植物[5]、锦鸡儿属植物[6]、四合木[7]、蒺藜科[8]等旱生植物,对同心圆枣的研究主要集中在同心圆枣的枣树栽培、植株离体培养[9]、病虫害调查防治[10]、促进枣树的生长发育和提高果实的品质等方面[11-12],对其的研究已经比较深入。同心圆枣在幼年期生长较为缓慢,但其具体原因没有研究报道,一般认为植物根茎叶的解剖结构与其抗旱性关系特征是植物抗旱的关键因素之一,而同心圆枣营养器官的结构特征与其抗旱性的关系尚未见报道,因此,拟对同心圆枣的结构特点与抗旱相关性进行研究。拟解决以下科学问题:1)揭示同心圆枣营养器官解剖结构与抗旱性的关系,为同心圆枣抗旱机理的研究提供形态解剖学依据;2)采用隶属度函数值法综合评价不同树龄同心圆枣的抗旱性强弱,为同心圆枣的栽培管理和进一步的开发利用提供理论基础和科学依据。
1 材料与方法
1.1 供试材料
试验材料取自宁夏同心县王团镇枣园(东经105°59′,北纬36°51′),该地区干旱少雨,蒸发量大,属于典型的大陆性气候。取材时选择生长健壮、具典型代表性的不同树龄枣树各3株,于生长旺盛季节6月分别采集两年生、十年生和百年生同心圆枣营养器官的根、茎和叶。根从根尖往上5 cm~6 cm处,截取1 cm小段,用自来水冲洗干净,茎选用发育良好的当年生枝条截取约1 cm小段,叶片在主脉两侧选取1 cm×1 cm小方块片,并固定于FAA(90 mL 50%乙醇+4 mL甲醛+6 mL冰乙酸)中。
1.2 切片制作与解剖结构观察
采用常规的石蜡方法制作切片[13],采集不同年龄植株的根、茎、叶,分割成小块,以FAA固定液固定,经酒精洗涤脱水、透蜡、包埋,常规石蜡切片,切片厚度为8μm~10μm,铁矾-苏木精染色,并以番红-固绿双重染色对照,中性树胶封片,Leica-DMLB显微镜观察并照相。由于同心圆枣的根、茎质地坚硬,故制片时进行适当的软化处理。
1.3 数据分析与处理
将显微镜图像处理结果或视野中所观测的数据记录在Excel中,用SPSS 22.0做差异显著性分析(P<0.05)。
隶属函数值的计算:
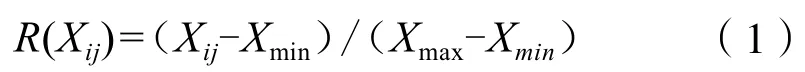
如果某一指标与抗旱能力成负相关,则用反隶属函数:

式中,R(Xij)为i年龄j指标抗旱性隶属函数值,Xij为i年龄j指标测定值,Xmax表示第j个综合指标的最大值,Xmin表示第j个综合指标的最小值。
采用于萍等[14]的方法利用Microsoft Excel 2010进行数据的灰色关联分析。
2 结果与分析
2.1 不同年龄同心圆枣根解剖结构分析
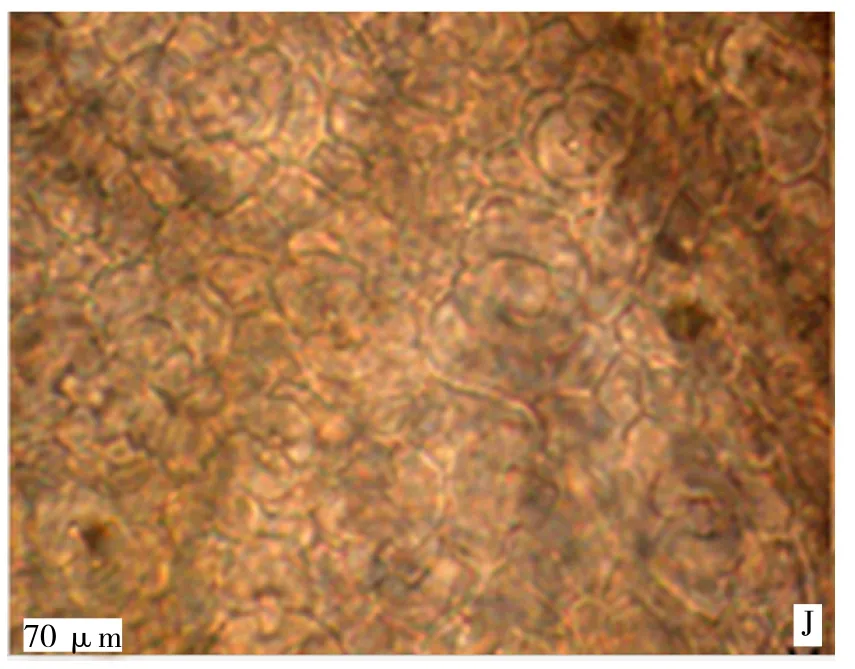
由图1可知,成熟根的横切面呈圆形,由周皮和次生维管组织组成。周皮由木栓层、木栓形成层和栓内层组成。最外的是木栓层,由3~5层细胞组成,其外侧的2~4层细胞呈褐色,因内部组织的增加而受压破坏和剥落,内侧1~2层扁平状细胞排列紧密;木栓形成层为一层扁平细胞,细胞壁较薄;栓内层为2~3层扁平细胞,细胞较小,排列紧密。次生维管组织由次生韧皮部、维管形成层和次生木质部组成。初生韧皮部已被破坏,经过次生生长形成次生韧皮部,由筛管、伴胞、韧皮纤维、韧皮部薄壁细胞和韧皮射线组成,韧皮部薄壁细胞中分布有大量的黏液细胞。

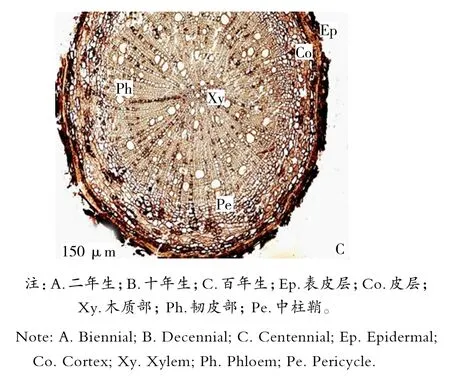
图1 同心圆枣根横切解剖结构Figure 1 Cross section of root in Zizyphus jujube Mill
由表1可知,不同年龄的同心圆枣根解剖特征值在木质部宽、木栓层厚度和导管直径等方面均存在着显著性差异,而木质部半径之间没有显著差异性,约为0.62μm;形成层由一层扁形细胞组成,呈棕黄色,细胞排列整齐。木质部由初生木质部和次生木质部组成,初生木质部二原型,在根的中央被保存下来;次生木质部位于初生木质部的外方,由导管、木纤维和木薄壁细胞以及木射线组成。木质部的宽度占根横切面半径的63%(表1),导管分子的直径达40.67μm。木质部宽和根半径的特征值趋势一致,表现出十年生的同心圆枣高于百年生和二年生;木栓层厚度则是同心圆枣百年生的最大,其次是十年生的,最后是二年生的,同心圆枣二年生的占百年生的71.5%;而导管直径则恰好与木栓层厚度相反,同心圆枣二年生最厚,同心圆枣十年生次之,同心圆枣百年生最薄,同心圆枣百年生导管直径约是同心圆枣二年生的0.44倍。
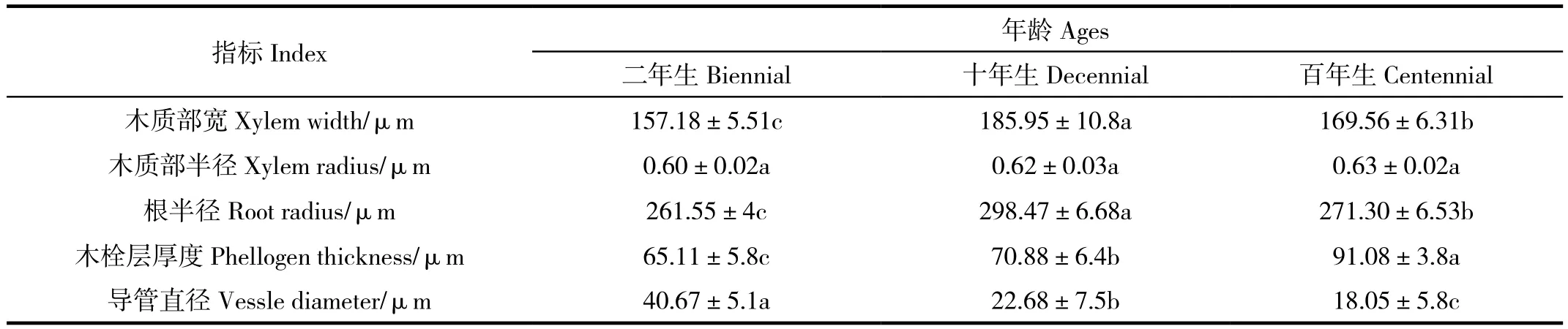
表1 不同树龄同心圆枣根的解剖学特征Table 1 Anatomical characters of roots in Zizyphus jujube at different ages
2.2 不同年龄同心圆枣茎解剖结构分析
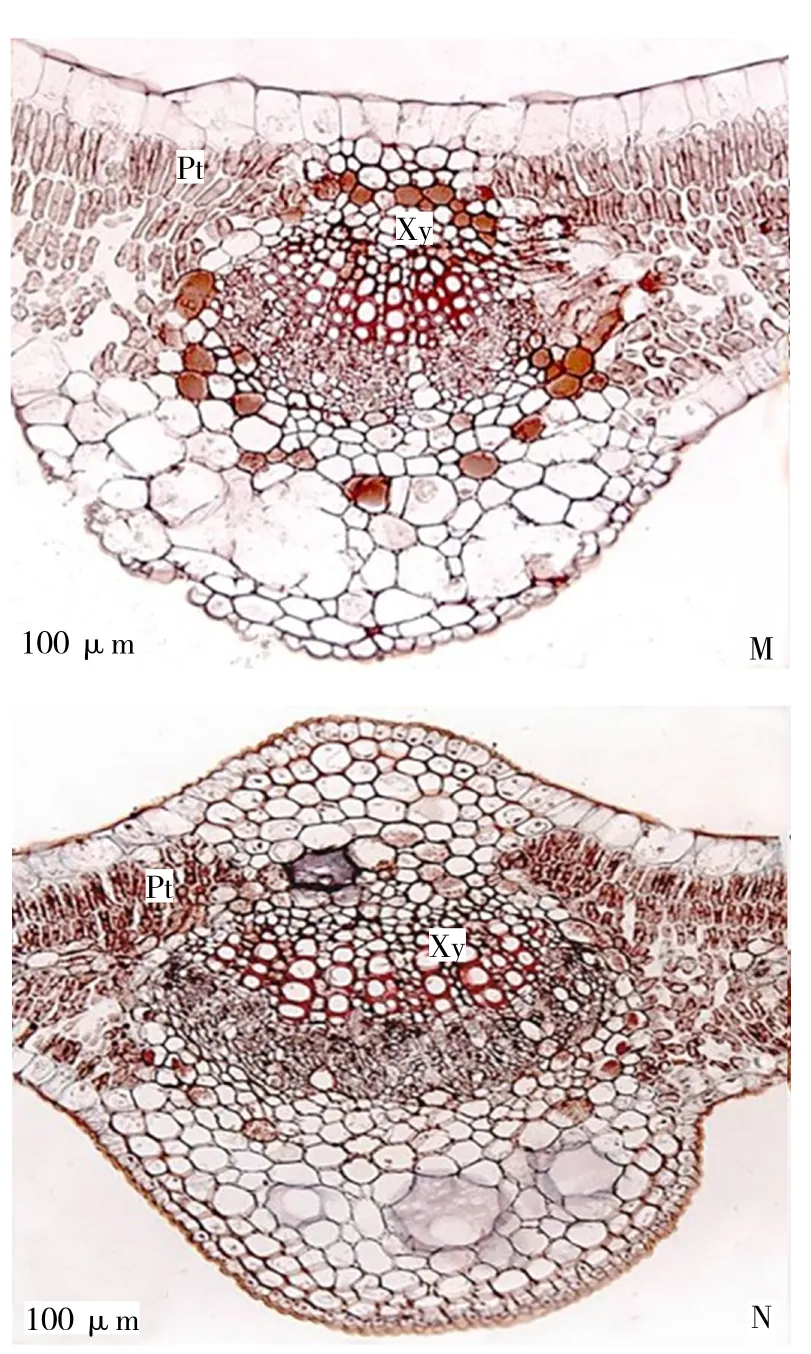
由图2可知,茎的横切面呈近圆形,具有明显的双子叶植物茎的结构特点,茎的初生结构自外向内由表皮、皮层、维管柱和髓构成;二年生茎在次生生长中形成周皮和次生维管柱。在老茎中,表皮被周皮代替,木栓层为深棕色,由多层厚壁组成;木栓形成层和栓内层。皮层由10~12层薄壁细胞组成,表皮内第1~5层细胞排列紧密,细胞壁较厚,且分布有一定数量的黏液细胞;第6~12层细胞较大,细胞排列较疏松,存在大量的细胞间隙。皮层在后期的次生生长过程中受内部组织的挤压而破坏。次生维管组织连接成环状,由次生韧皮部、维管形成层和次生木质部组成。其中韧皮部由筛管、伴胞和韧皮薄壁组织构成,韧皮部薄壁细胞中分布了少量的黏液细胞。
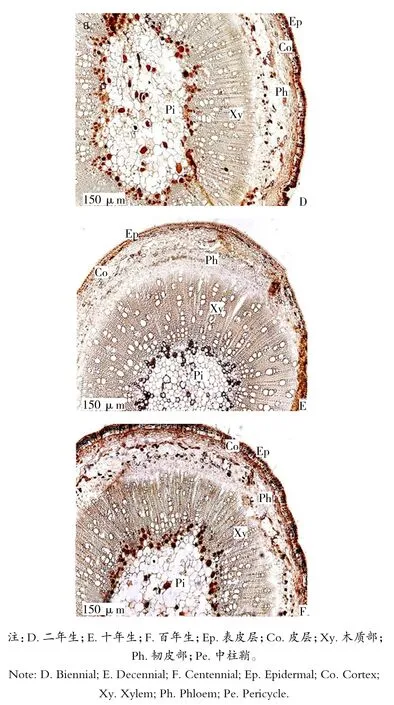
图2 同心圆枣茎横切解剖结构Figure 2 Cross section of stem in Zizyphus jujube Mill
由表2可知,不同年龄同心圆枣茎半径之间存在着显著性差异,木质部的宽度占茎横切面半径的41%,木质部导管分子呈放射状排列,木质部中分布密度为56个/平方毫米。维管射线由2~3层细胞组成,呈放射状排列。髓部较发达,由大量薄壁细胞构成,细胞间隙大,其总面积占茎横切面总面积的33.4%,靠近木质部内侧的髓部分布有一层黏液细胞。
表2 不同树龄同心圆枣茎的解剖学特征Table 2 Anatomical characters of stems in Zizyphus jujube at different ages
2.3 不同年龄同心圆枣叶解剖结构分析
通过图3可知,同心圆枣叶片为等面叶,无海绵组织的分化,叶片较厚,约150 μm左右,叶两面无毛,成熟叶片由表皮、叶肉和叶脉组成。上下表皮各一层细胞,大型表皮细胞呈长方形或方形,排列紧密,上表皮细胞较下表皮大,约34 μm;表皮外有发达的角质层;气孔仅分布于下表皮,气孔较小且分布密集(见图4);叶肉组织发达,由5~6层栅栏组织薄壁细胞组成,栅栏组织厚度约124.3 μm,占叶片厚度的73.5%。叶子近轴面2~3层长柱状细胞排列紧密,胞间隙小,而远轴面3层短柱状细胞排列相对疏松;叶脉维管束发达,并被圆形、不规则薄壁细胞组成的发达维管束鞘所包围,其背腹面均有厚壁细胞分布;由图5可知,叶片主脉中的薄壁组织分布有黏液细胞围绕维管束排列,中肋韧皮部中分布有簇晶细胞。同心圆枣百年老树主脉明显多且发达,厚度也大于十年生和二年生的。

图3 同心圆枣叶片横切解剖结构Figure 3 Anatomical structure of leaf cross section of Zizyphus jujube Mill

图4 氢氧化钠透明法对同心圆枣叶片气孔的观察Figure 4 Showing the distribution of stoma in Zizyphus jujube Mill.

图5 同心圆枣叶片主脉横切解剖结构Figure 5 Showing the anatomical structure of the medial vein in Zizyphus jujube Mill
通过表3可以看出不同年龄同心圆枣叶片解剖特征值之间基本都存在显著差异性,栅栏组织厚度和主脉直径的变化是由同心圆枣百年老树向同心圆枣二年生降低的过程,同心圆枣老树的栅栏组织厚度比同心圆枣二年生高12.54%,同心圆枣老树的主脉直径比同心圆枣二年生高出近40%;上表皮厚度在同心圆枣二年生和同心圆枣十年生之间差异不显著,同心圆枣老树表现差异显著;下表皮厚度随着枣树年龄的增长而降低,呈显著差异性,同心圆枣老树是同心圆枣二年生的39.5%;角质层厚度之间差异不显著,基本没变化;叶片厚度总体呈升高的趋势,在同心圆枣二年生和同心圆枣十年生之间有显著差异性,与同心圆枣老树差异不显著,但同心圆枣二年生叶片厚度是同心圆枣十年生的86.3%,是同心圆枣老树的79.7%;同心圆枣老树的导管直径稍粗于同心圆枣十年生和同心圆枣二年生的;黏液细胞直径在不同年龄的枣树中差异不显著;同心圆枣老树的气孔数量要明显少于同心圆枣二年生和同心圆枣十年生的,比同心圆枣二年生的约少18.5%。

表3 不同树龄同心圆枣叶子的解剖学特征Table 3 Anatomical characters of leaves in Zizyphus jujube at different ages
2.4 不同年龄同心圆枣营养器官解剖结构综合评价
从表1~3的数据可知,不同年龄的不同指标间变化不一致,说明在分析不同年龄同心圆枣的抗旱性时,不能根据一项指标的优劣判断其抗旱性。因此,本研究从根、茎、叶中挑选出差异性显著的指标(见表4),标准化后采用隶属函数值法(根导管直径采用反隶属函数),对不同年龄的同心圆枣具有显著差异性的指标进行综合评价,取平均值。从结果可以看出,不同年龄的同心圆枣的抗旱性强弱不同,二年生同心圆枣树的隶属函数平均值为0.10,十年生的为0.72,百年老树的为0.74,抗旱性大小排序为同心圆枣百年老树>同心圆枣十年生>同心圆枣二年生,且同心圆枣百年老树和十年生的抗旱性远大于二年生的同心圆枣。

表4 不同年龄抗旱性综合评价Table 4 Comprehensive evaluation of drought resistance at different ages
2.5 同心圆枣抗旱性的灰色关联分析
将不同树龄同心圆枣的抗旱性指数及10个指标视为一个整体,即灰色系统。原始数据通过计算机软件标准化处理后,计算各指标与抗旱性指数的关联系数,然后分别求出各指标与抗旱性指数的关联度,并按关联度大小排列出关联序。
将不同树龄同心圆枣的抗旱性指数作为参考数据列(母序列,隶属函数平均值标准化得出),记为X0=(0.192 308,1.384 615,1.423 077);选取营养器官结构特征的10项抗旱指标为比较数列(子序列),记为(i=1,2,…,9),X1=(1.053 146,0.995 456,1.046 385),X2=(1.752 452,1.598 624,1.662 681),…,X10=(2.389 915,2.535 724,2.688 527),分别求出各指标变化率(Xi)与(X0)的关联度,并按照关联度大小进行排序(见表5)。
依照关联分析原则,关联度愈大,表明参考各指标与抗旱性的关系愈紧密,反之,关系较远。因而,关联度大小可表明某一项指标对抗旱的敏感性。从表5可以看出,植物的抗旱性与各指标的关联序为茎直径(X5)>叶主脉直径(X10)>根半径(X2)>根木质部宽(X1)>叶片栅栏组织厚度(X9)>根木栓层厚度(X3)>茎导管密度(X7)>根导管直径(X4) >叶下表皮厚度(X8)>茎角质层厚度(X6)。
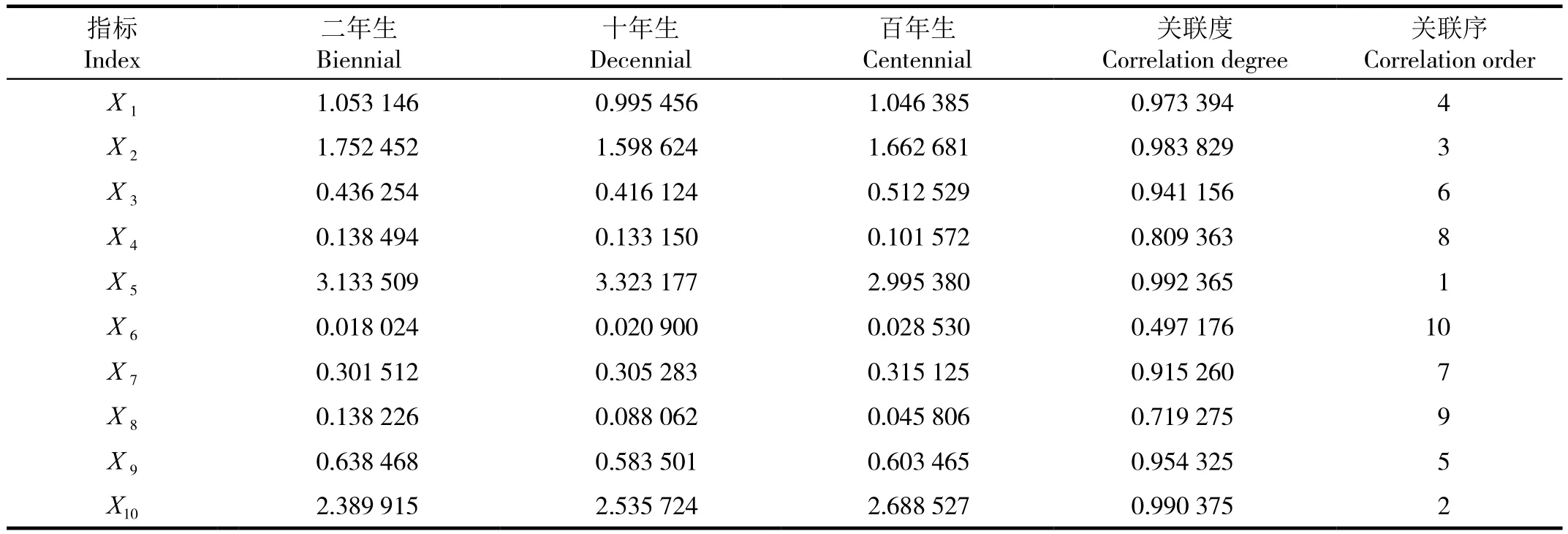
表5 不同树龄同心圆枣抗旱性与各种指标的关联系数、关联度及关联序Table 5 The indexes of drought resestance and their correlative coefficient, correlative degree and correlation order
3 讨论与结论
3.1 根解剖结构与抗旱性
同心圆枣木栓形成层在生长期内不断分生加厚,向外形成木栓层,向内形成栓内层,栓内层越发达,表明植物的物质储备能力越强[15]。栓内层具有减少植株体内水分外散的作用,是对旱生环境的适应[16]。本研究中百年老树的木栓层厚度和木质部厚度明显高于十年生和二年生的,说明同心圆枣老树通过根部的变化适应环境,增加其抗旱性,根木质部越厚,韧皮部越发达,运输物质的能力越强,可以吸收更多的物质,则抗旱性越强,这与陈小红等[17]对川西高原4种高山海棠的营养器官解剖结构中的研究结果相一致,在植物的吸收根中,皮层厚度的变化可用于评估植物对环境变化的适应性[18]。一种观点认为导管孔径越大,说明运输水分的阻力越小,输水效率越高[15],也有学者认为,土壤水分比较缺乏环境下的根导管较大容易造成导管空腔和栓塞,不利于形成根压向上长距离运输[19]。李鲁华等[20]在小麦根系与土壤水分胁迫关系的研究进展中认为较小直径的导管可以减少在干旱环境中开花或敏感时期使用的土壤水的消耗,从而提高产量和水的利用率,这与本研究的结果一致,在今后研究中,要特别注意果树不同发育时期根系形态的变化。同心圆枣根的韧皮部中存在黏液细胞,黄振英等[21]认为韧皮部中黏液细胞内存在的黏液物质为树胶,具有保水能力,为周围的细胞提供一个较湿的小环境,是旱生植物所具有的抗旱特征。同心圆枣根木质部导管分子的直径达40.67μm,强大的疏导系统为水分和矿物质从植物根部输往各同化器官提供了保证。
3.2 茎解剖结构与抗旱性
一般认为:茎的表皮和角质层形成了保护结构,以防止在贮存和运输过程中水分流失,是对干旱环境长期适应的结果[22-23]。表皮可以保护木材和韧皮部免受生物和非生物胁迫[24],皮层外侧分布的大量黏液细胞,能增加细胞液浓度,改变细胞渗透压,提高了吸水和贮存水分的能力,增强抗旱能力;密集的导管细胞提高了枣树在干旱环境中的输送效率,发达的髓部能贮藏降雨时期多余的水分,并且髓部外侧的一层黏液细胞能起到保水的作用[21],百年老树的黏液细胞直径要大于十年生和二年生的,相比可以更好地保存水分。同心圆枣老树茎的角质层厚度和木质部宽度都大于十年生和二年生的,这与刘新雨等[25]关于菊芋的研究结果一致,即植株通过增加角质层厚度和木质部宽度来抵御干旱。不同年龄的同心圆枣茎的解剖结构及参数值在一定程度上反映了其具有良好的抗旱性。
3.3 叶片解剖结构与抗旱性
解剖结果发现同心圆枣叶片的解剖结构为等面叶,与王中英等[26]关于枣树的组织解剖构造中的结果一致。随着树木高度或年龄的增加,叶片会相应地发生一系列的变化,如比叶重增加、叶面积减小和角质层厚度增加等[27]。植物叶片角质层的加厚自然可以抑制蒸腾失水[28],较大的表皮细胞具有贮水作用,对于增强水分的调节能力有一定意义[29]。植物叶片中的海绵组织越厚,叶片疏松度越大,说明叶肉细胞排列越疏松,则会使叶片的抗旱性越差[30-31],而在观察同心圆枣的叶片解剖结构中没有发现海绵组织,这与曹娟云等[32]对四种枣叶片的解剖学研究结果一致,说明同心圆枣的确具有较强的抗旱性。植物叶片越厚,表明其储水能力越强,本研究中同心圆枣百年生老树的叶片厚度要明显高于十年生和二年生的,这与潘昕等[30]对25种青藏高原灌木叶片解剖结构与抗旱性中的研究结果相近。叶片表皮细胞能有效避免强烈光照对叶肉细胞的灼伤,也可以有效利用衍射光进行光合作用,使植物利用光能的效率更高,是对干旱的高度适应性,同心圆枣具有这一特征;同心圆枣中含晶细胞的出现则可以降低积累的有害物质浓度,形成一个相对稳定的环境,以积极方式适应干旱环境;叶片主脉主要起输导和支持的作用,发达的中脉不仅能很好地输送水分、养分而且有保水、贮水的作用,主脉中的薄壁组织分布的黏液细胞能够使叶片中的水分不易散失[33],这种细胞分布形式能在干旱季节使周围的细胞形成湿润的小环境,这一点对中部干旱带的植物是十分重要的。
3.4 植株年龄与抗旱性
李吉跃[34]对太行山区的造林树种研究发现,同一树种的保水力在不同年龄间存在差异,且不同的树种随年龄的变化趋势不同。李淑英等[35]研究发现,二年生的常春藤的抗旱性大于一年生的,可见植物的抗旱性与其年龄存在一定的相关性。又有研究发现干旱区油松的保水力随着年龄的增加而下降,而侧柏则是有所增强,阔叶树种有的随着年龄增加抗旱性在增强,如白蜡,元宝枫;有的变化则不大,如刺槐和紫穗槐;有的反而增强,如黄栌和沙棘[34]。各树种各年龄间的抗旱性是否有显著性差异,并无明确判断,但足以说明不同年龄树木的抗旱性增加或降低,存在着一定差异,这种变化将会影响它们在长期干旱条件下的生存能力[36]并及时完成对环境变化的响应。
从解剖结构可以看出,不同年龄的同心圆枣的根茎叶等营养器官都具有一定的抗旱性特征。同心圆枣抗旱机理是以尽可能地保存水分、减少叶片失水、提高光合效率占主要地位,在水分有限的状况下,最大限度吸收水分和储藏水分来调节体内的水分平衡,因此同心圆枣耐旱性是以忍耐性的方式适应干旱严酷环境。隶属函数值分析表明同心圆枣百年老树的抗旱性相对更强,但十年生的同心圆枣树的抗旱性也相当强。因此,在生产中选择强抗旱性的同时还应该结合枣树的产量进行综合研判,以获得最大效益。灰色关联度分析结果表明茎直径(X5)、叶主脉直径(X10)、根半径(X2)、根木质部宽(X1)和叶片栅栏组织厚度(X9)等可以作为同心圆枣抗旱解剖结构的主要鉴定指标,以后也可以通过鉴定这些指标的大小判断其他植物抗旱性强弱。
猜你喜欢
作物研究(2022年4期)2022-09-21
华人时刊(2022年3期)2022-04-26
黄河之声(2022年1期)2022-03-16
中国糖料(2021年3期)2021-07-13
黄河之声(2021年21期)2021-03-22
植物研究(2021年2期)2021-02-26
种子(2020年11期)2020-12-05
华人时刊(2020年19期)2020-11-17
天津农业科学(2015年11期)2015-12-03
安徽农学通报(2014年7期)2014-04-29
