
异甾体生物碱类化合物及药理活性研究进展
2022-02-19 13:39王安汶刘玉明
天然产物研究与开发 2022年1期
王安汶,刘玉明,2*
1天津理工大学化学化工学院;2天津市药物靶向与生物成像重点实验室,天津 300384
异甾体生物碱(isosteroidal alkaloids)是属于甾体生物碱中一类结构多样并具广泛药理活性的生物碱,主要分布于百合科贝母属Fritillaria和藜芦属Veratrum植物中,为两属植物发挥药理活性的主要成分。异甾体生物碱结构小众,数量不多,但是其独特的结构及药理活性持续引起国内外学者关注。鉴于2006年前有关天然来源异甾体生物碱类研究已有总结[1,2],因此本文综述了十五年间(2006~2020年)新发现异甾体生物碱(共80个)的植物来源、化学结构及药理活性,以期为异甾体生物碱类成分的进一步研发提供科学依据。
1 异甾体生物碱分类
异甾体生物碱通常具有C27骨架体系,基本骨架结构具有5~6个碳环,其特征环型为C-nor-D-homo-[14(13→12)-abeo]环系,其中包含哌啶环单元。其化学结构复杂多变,可因羟基、羰基、双键等取代基团的位置和个数或构型、20或25位上的甲基构型、是否N-氧化物、D/E等环顺反异构、环系开裂或新环系形成等因素而使活性产生明显差别。现根据骨架结构主要分为西藜芦碱型(cevanine type)、藜芦胺型(veratramine type)和介藜芦型(jervine type)三类(见图1)。
1.1 Cevanine(A)型生物碱
该类型生物碱的骨架为六个环共同骈合而成,其中A/B/D环为六元环,C环为五元环,E环或F环的18-、22-和26-位碳与同一氮原子相连。Cevanine型生物碱在贝母属植物和藜芦属植物中所占比例为最多,近十五年间新发现的cevanine型生物碱共有35种,其化学结构见图2,名称及来源见表1。
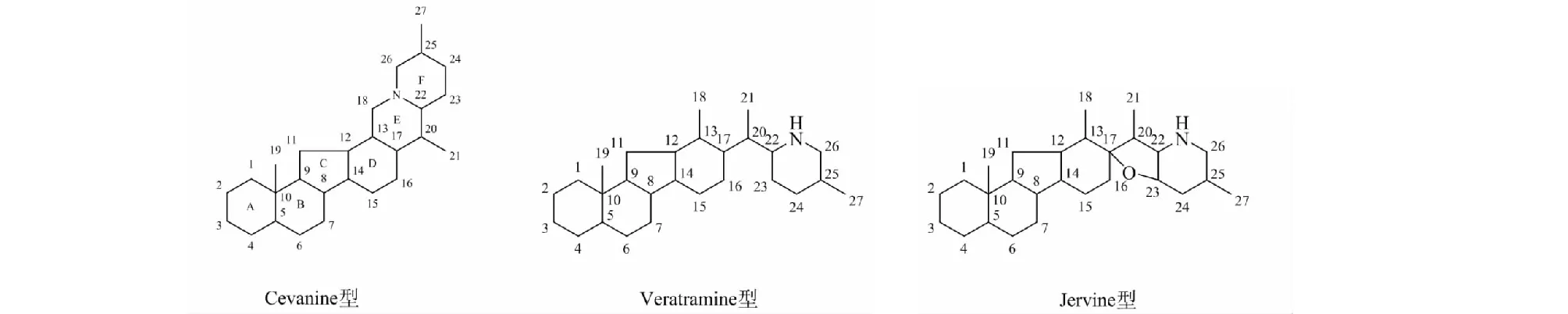
图1 异甾体生物碱骨架结构Fig.1 Basic skeletons of isosteroidal alkaloids

图2 新发现cevanine型生物碱的化学结构Fig.2 Structures of new cevanine-type alkaloids

表1 新发现cevanine型生物碱的名称及来源
1.2 Veratramine(B)型生物碱
Veratramine类型生物碱为五环架构,与cevanine型异甾体生物碱的区别在于其E环的C-N键断裂。在新发现的异甾体生物碱中所占比例最低,有20种,其化学结构见图3,名称及来源见表2。
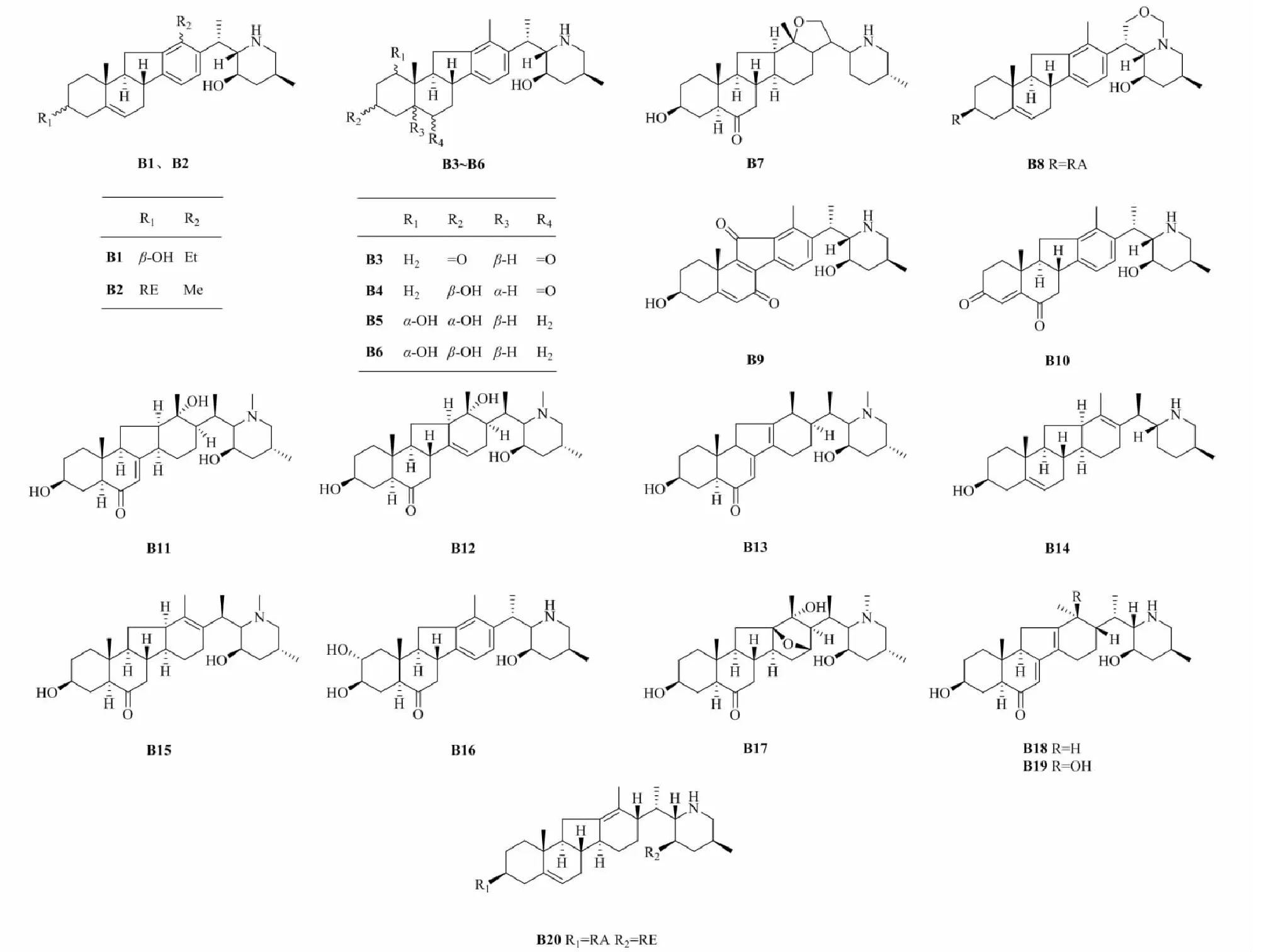
图3 新发现veratramine型生物碱的化学结构Fig.3 Structures of new veratramine-type alkaloids
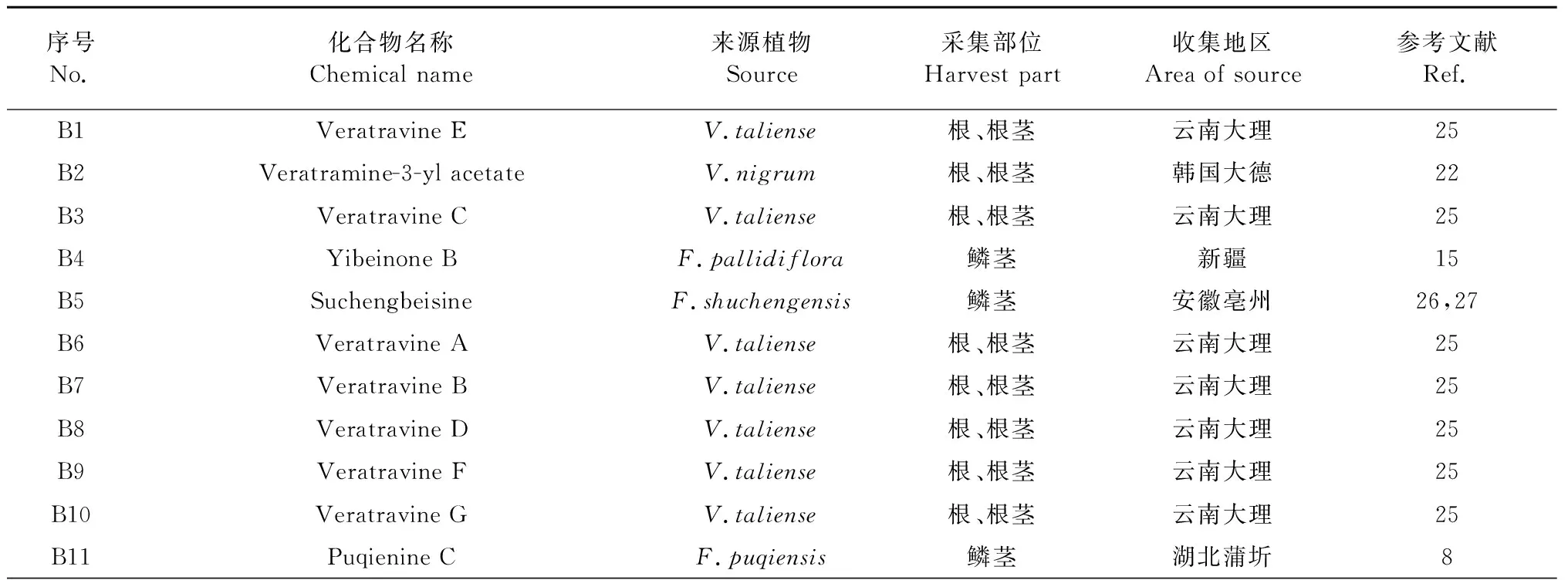
表2 新发现veratramine型生物碱的名称及来源

续表2(Continued Tab.2)
1.3 Jervine(C)型生物碱
Jervine型生物碱骨架与cevanine型骨架相似,区别为E环为四氢呋喃环。在新发现的异甾体生物碱中,数量少于cevanine型结构,为25种,其化学结构见图4,名称及来源见表3。

图4 新发现jervine型生物碱的化学结构Fig.4 Structures of new jervine-type alkaloids
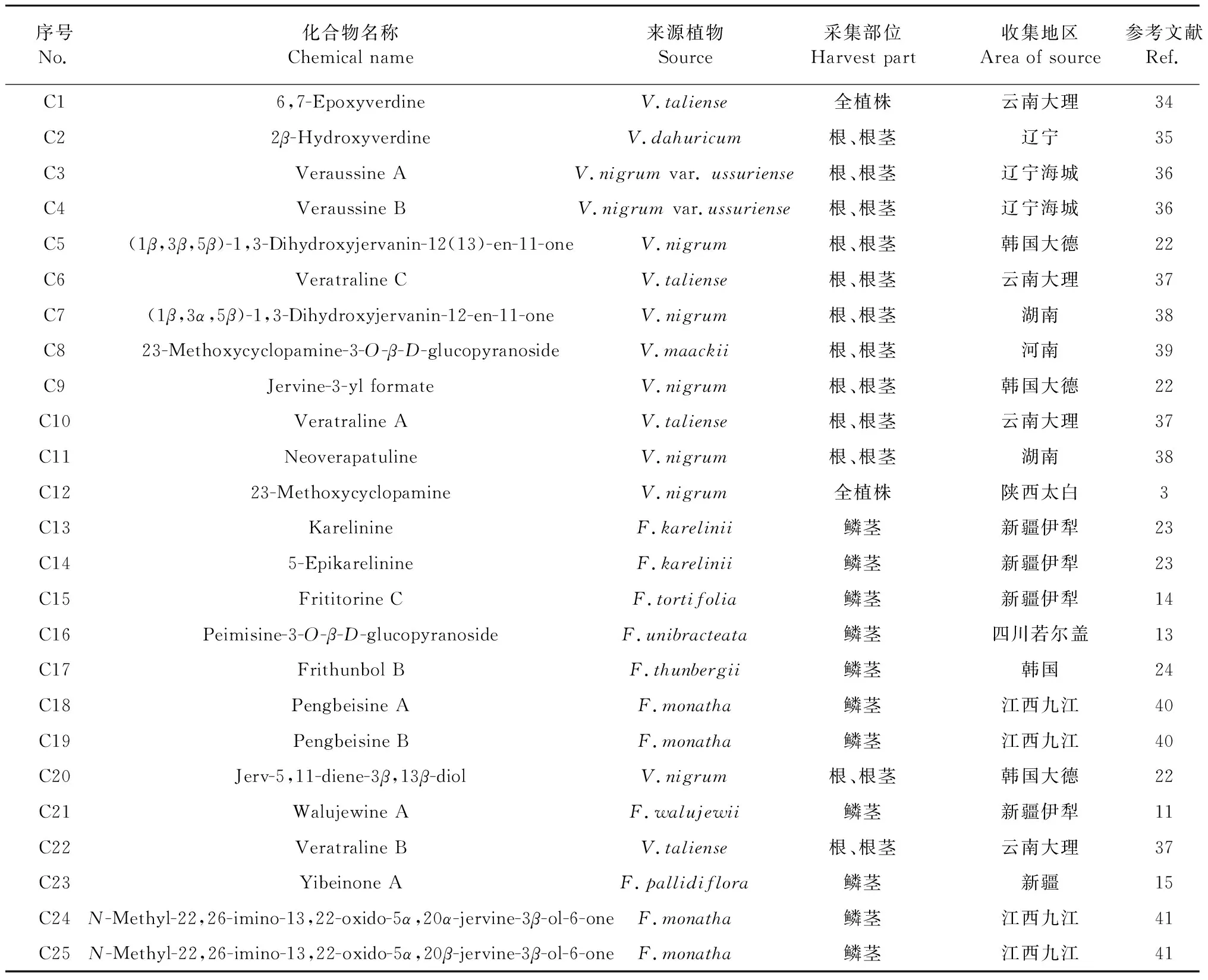
表3 新发现jervine型生物碱的名称及来源
植物中异甾体生物碱的含量受多种因素影响,包括温度、产地、部位和种类等。研究表明暗紫贝母随着海拔升高,生物碱含量随之上升,温度的降低有利于植物体内生物碱含量的积累,但超过一定范围后不利于植物的生长[42]。除温度外,不同产地同种植物中生物碱含量亦有不同,在六个浙贝母产区中,磐安(0.53%)、缙云(0.52%)、象山(0.52%)产区生物碱含量较高,而东阳(0.44%)、鄞州(0.41%)、舟山(0.38%)产区较低,且差距达到0.15%[43]。此外,浙贝母不同部位含生物碱总量也有差别,Chen等[44]测定和比较了浙贝母花、茎、鳞茎的总生物碱含量,结果显示花为0.182 %,茎为0.047 %,鳞茎为0.341 %,这也与大多采用贝母鳞茎作为药用部位相符。还有不同种类贝母所含生物碱总量有着极大的差异,相关研究[45]比较了平贝母、太白贝母、松贝、青贝、炉贝的总生物碱含量,测定结果显示太白贝母(野生)>平贝母>青贝>松贝>炉贝>太白贝母(栽培),其中太白贝母的野生品种(0.221 8%)与栽培品种(0.057 6%)的差距近四倍。总之,植物中异甾体生物碱含量稀少,且受内部因素与外部因素影响而差异不小,了解这种差异对品种选育和栽培研究极具价值。目前未见对藜芦属植物的相关研究报道。
2 异甾体生物碱的化学合成
部分异甾体生物碱对肿瘤干细胞有抑制作用,其中最著名的当为Hedgehog(Hh)信号途径传导抑制剂环巴胺。天然异甾体生物碱几乎都来源于贝母属和藜芦属植物,而从中提取分离困难重重,得率不尽如人意,这使得异甾体生物碱的合成成为必然。异甾体生物碱本身的C-nor-D-homo-[14(13→12)-abeo]环型对合成有着相当大的难度,藜芦属植物中异甾体生物碱的生源途径决定了工业合成几乎难以做到[46]。目前合成研究最多的是环巴胺及其相关衍生物,早前Giannis等[47]便使用去氢表雄酮为原料合成环巴胺,反应过程共有20步,接近1%的总产率。Mousavizadeh等[48]重复了该过程,得到了相同的结果。此外Mousavizadeh及其同事还使用了路易斯酸介导的Nazarov Cyclization法,用以[C-nor-D-homo]-生物碱的一般合成以及相关转化方法(图5)。值得注意的是,反应最后一步转化使用的路易斯酸仅TiCl4取得成功,其他如FeCl3、Cu(OTf)2、Sc(OTf)3、AlCl3、BF3·OEt2及Brφnsted酸(TsOH、TFA)均以失败告终。在此之前,Heretsch等[49]便介绍了另一种合成[C-nor-D-homo]-生物碱的方法(图6),区别在于使用Schönecker’s C-H活化/羟化手段脱去原料中的C-12羟基,重排形成C-nor-D-homo-环结构,并在C-12与C-13间形成碳碳双键,其他相似物质的合成结果表明该方法同样适用。C/D环的构建是异甾体生物碱合成的关键,Heretsch的方法针对具有特定结构的底物,而Mousavizadeh等给出的转化方法采用简单的双环结构,在异甾体生物碱的合成上具有一定意义。环巴胺的合成也可以从已知相似结构的生物碱修饰得到,如将贝母辛转化为环巴胺(图7),其过程简单,并且反应条件温和,产率高,转化率可达93%,原料获取较为容易[50]。还有研究报道[51]以ketone(±)-4化合物为原料,合成去氢表雄酮,这不仅为环巴胺的合成原料提供了一种解决方案,也为环巴胺的工艺路线提供了新的思路。
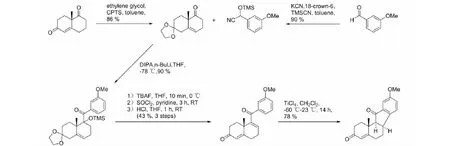
图5 [C-nor-D-homo]-生物碱的合成Fig.5 Synthesis of a C-nor-D-homo-steroid
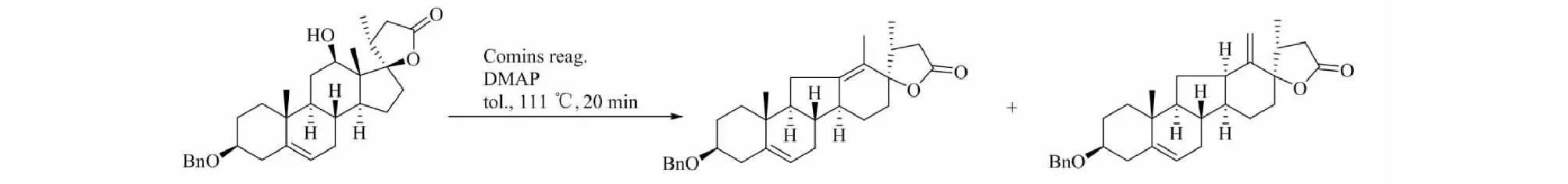
图6 C-nor-D-homo-环合成Fig.6 C-nor-D-homo-cyclosynthesis
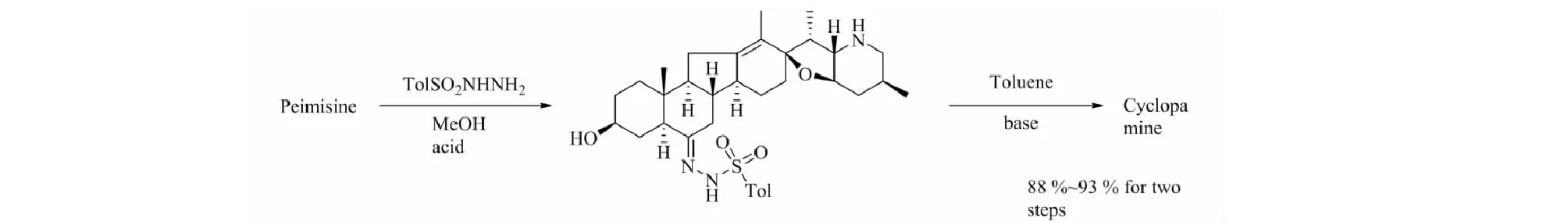
图7 贝母辛转化为环巴胺Fig.7 The conversion of peimisine to cyclopamine
Hh信号途径被发现后,环巴胺一直是备受关注的抗癌候选物,而对其相似化合物亦多有研究。环巴胺的缺点是酸不耐受、易水解成有毒的藜芦胺。鉴于此Rabe等[52]设计了对Hh信号途径有着类似作用活性的类似结构,与环巴胺不同的是F环采用的是哌啶结构,并去除了C-21甲基,同样经由去氢表雄酮原料合成得到。另为寻找酸耐受的环巴胺类似物,在环巴胺C-13位进行改性研究,其合成的类似物bis-exo-cyclopamine在C-13和C-25位均改为碳碳双键,其余与环巴胺相同,在pH=1的酸性条件下,24 h后结构依然稳定,总产率可达18%[53]。
3 药理活性
3.1 抗炎作用
炎症症状程度分轻度与重度,严重的炎症反应会引起病理改变,给人带来红肿热痛等痛苦,甚至造成脏器功能受损和破坏,故关于炎症的治疗研究意义重大。NF-κB是已知炎症相关的重要因素,调控着不同促炎蛋白的表达。Zhou等[54]使用基于UPLC-Q/TOF MS的综合策略,通过PCA和ANN分析方法对五种贝母属植物(浙贝、松贝、平贝、青贝、炉贝)的生物碱进行研究,发现浙贝中生物碱可有效抑制炎症反应,在高剂量10 mg/mL和中剂量3 mg/mL的情况下,能够有效抑制TNF-α诱导的HEK 293细胞中的NF-κB信号通路的表达,且剂量越高,效果越明显,并优于其他贝母组。其分析结果表明cevanine型生物碱C-6羟基、C-20羟基和C-6羰基是抑制NF-κB信号通路的有效基团。Wu等[55]对RAW 264.7巨噬细胞促炎性介质的研究发现,川贝中西贝素和贝母素乙能够抑制NF-κB信号通路,从而达到抑制NO的产生和促炎因子的过表达,其机制在于两化合物可以通过抑制IκBα磷酸化来抑制NF-κB的核转录和转录活性,实验表明两化合物活性在0~150 μM浓度范围内呈正量效关系。值得关注的是西贝素和贝母素乙两者都有C-6羰基和C-20羟基取代,差别在于前者为D/E顺式构型,后者是反式构型,但在抑制NO、TNF-α、IL-1β、COX-2及IκBα等促炎因子活性上无明显差异。另在脂多糖(LPS)诱导的RAW 264.7巨噬细胞研究[56]表明:五种异甾体生物碱(imperialine、verticinone、verticine、delavine和peimisine)均能减少NO、TNF-α和IL-6产生,抑制TNF-α和IL-6的mRNA转录,同时这五种异甾体生物碱抗炎机制还与抑制丝裂素活化蛋白激酶(MAPK)信号通路磷酸化有关。其中cevanine型生物碱因同时具有C-6羰/羟基和C-20羟基而具有较好的抗炎效果,且强于jervine型生物碱,同样其D/E顺反异构对抗炎作用没有明显影响。
3.2 镇痛作用
镇痛药能为患者缓释痛苦、减轻疼痛,是临床常用药物,但含异甾体生物碱成分的止痛药少见。近年研究发现异甾体生物碱具有良好的止痛效果。Li等[25,37]对藜芦属植物大理藜芦中提取出的异甾体生物碱进行了抗炎止痛研究,所得到的异甾体生物碱均显示出良好的止痛作用,发现五种异甾体生物碱的镇痛效果强于常用药哌替啶。其中jervine型生物碱的镇痛效果均好于哌替啶,C-1和C-3羟基取代可能对抑制活性有较大影响,如失去两个羟基基团的dihydrojervine镇痛活性低于其他同类生物碱,不过C-6位上羟基取代后会使活性减弱;而在veratramine型生物碱中C-3取代基种类及构型、C-6羰基取代及其共轭体系可能起主要作用。
3.3 降压作用
血管紧张素转化酶(ACE)普遍存在于人身体多个部位,位于肺部的ACE含量丰富且活性最高,其可强烈收缩血管,增高血压。An等[57]评估蒲圻贝母中异甾体生物碱降压作用时,发现veratramine型生物碱对ACE具有一定的抑制作用,其中C-6位羰基与双键C-7(8)及C-14(12)形成的长共轭体系应是发挥ACE抑制作用的关键因素。另有研究[58]表明实验中jervine型生物碱均有一定的降压效果,A环和B环中羟基较多的verdine生物碱显示出更强的降压作用,尤其是B环6位羟基存在时降压效果显著;而相比较cevanine型生物碱的降压活性不明显。
3.4 平喘作用
哮喘是全球人群饱受折磨的呼吸道疾病,可由多种因素引起,病情顽固且难以根除,严重时可造成生命危险,如何有效改善和解决哮喘疾病是目前世界性疾病难题之一。Wnt/β-catenin信号通路是参与哮喘发生的重要通路,Wnt信号通路的配体Wnt5a和Wnt7b在调控气管及肺部平滑肌中必不可少,而β-catenin在细胞的生长、增殖以及稳态维持起着重要作用。当支气管过度收缩或拉伸时,会激活β-catenin进而促进气道上皮细胞增殖、分化和细胞外基质生产,使组织修复、纤维化以及哮喘气道重塑[59]。近年发现伊贝母内异甾体生物碱A22和A23均有良好的平喘作用,其平喘能力强弱为氨茶碱(pA2=7.32±0.08;EC50=0.05±0.00 μM)>A22(6.19±0.02;0.65±0.02 μM)>A23(5.58±0.09;3.00±0.68 μM)>硝苯地平(5.22±0.08;6.50±1.00 μM),其可能的平喘作用机制即是通过抑制Wnt5a或者Wnt7b的蛋白表达,从而抑制β-catenin激活,进而阻止了哮喘气道炎症和气道重塑的发生发展[15,59]。还有托里贝母中新发现的异甾体生物碱A20对ACE诱导的气管收缩表现出明显的舒张作用,其pA2值为5.97±0.92 μM,EC50值为3.40±4.90 μM,舒张能力与氨茶碱相当[14]。另外Cui等[60]采用网络药理学结合分子对接的研究方法,发现浙贝母花中异甾体生物碱贝母素甲和贝母素乙可通过作用于PTGS2、AKT1等靶点而抑制呼吸道炎症,减少黏液分泌,抑制气管重塑。以上表明,cevanine型生物碱应是异甾体生物碱中具有抗哮喘功效的主要结构类型,C-6位羰基以及C-20位羟基结构应是发挥抗哮喘活性不可或缺的存在,而3位羟基的糖苷化会使得活性减弱,氮氧化物的存在也会削弱其平喘作用。
3.5 抗血栓作用
Song等[61]发现黑藜芦总生物碱有着不俗的抗血栓能力,其中分出的藜芦酰基棋盘花胺是唯一在各个剂量都强于总生物碱的化合物,可能为总生物碱中起抗血栓活性的主要成分。值得注意的是,cevanine型生物碱都表现出或强或弱的抗血栓活性。从初步构效关系来看,cevanine型骨架可能是抗血栓活性所必需,而C-3位酯基的取代能够增强其抗血栓作用。鉴于血栓与血小板的血管聚集程度相关联,Tang等[62]对兴安藜芦进行了研究,从中得到的五种cevanine型生物碱都表现出抑制血小板聚集活性,其中15-angyloylgermine生物碱在花生四烯酸诱导的培养基上显示了最强的抑制作用(200 μM时为100%,100 μM时为95.4%)。与之对应的veratramine型生物碱抑制率较低,仅介藜芦胺的抑制率略高。故cevanine型生物碱可能具有普遍的抗血栓活性,可进一步作为抗血栓药物的先导物开发,不过目前尚缺乏后续的药代动力学和临床数据。
3.6 胆碱酯酶抑制作用
乙酰胆碱酯酶(AChE)与丁酰胆碱酯酶(BChE)活性的增加是反映阿尔茨海默病(AD)的重要指标。AChE抑制剂是目前临床主要使用的抗AD药物,而对AD中后期来说BChE则是更有价值的药物作用靶标。近年研究[11]发现cevanine型生物碱对BChE的抑制活性强于AChE,不过对这两种酶抑制活性的强弱顺序一致;与之相反jervine型生物碱则为选择性AChE抑制剂。值得注意的是,cevanine型生物碱C-6位β羟基取代后的抑制活性强于α羟基取代,而C-20位羟基取代会导致其抑制活性消失。另外jervine型生物碱中N-αH取向、末端双键和C-12羟基的存在对AChE选择性抑制具有重要影响。再有研究表明[63]cevanine型生物碱hupeheninoside和chuanbeinone均为选择性BChE抑制剂(IC50=2.1 μM和0.7 μM),而C-20羟基取代的cevanine型生物碱均未表现出ChE抑制作用,N-甲基化后也极大地削弱了其抑制活性。比较特殊的是,sinpeinine A和chuanbeinone结构相近,其为cevanine型生物碱E/F环的顺反异构体,但结果表明chuanbeinone抑制活性远高于sinpeinine A,提示E/F顺式构型(22-βH)有利于胆碱酯酶抑制作用。
3.7 抗病毒作用
流感病毒可引起具高发病率和高死亡率的呼吸系统疾病。Kim等[64]评估了F.thunbergii提取物对流感病毒(H1N1)感染的治疗效果,体外研究结果表明:与oseltamivir对照相比(CC50:733 μg/mL;EC50:3.3 μg/mL),F.thunbergii提取物CC50值高达7 500 μg/mL,EC50为148.2 μg/mL,说明其具有中等抗流感H1N1病毒效果,且毒性很低。且在以网络药理学预测F.thunbergii治疗流感的潜在成分中,确立了两个化合物天竺葵色素和β-谷甾醇及其与流感相关的九个靶基因(BCL2、CASP3、HSP90AA1、ICAM1、JUN、NOS2、PPARG、PTGS1和PTGS2)之间的相互关系,建立了成分-流感等网络,揭示了F.thunbergii成分发挥流感治疗作用的内在联系[65]。除流感病毒外,Li等[21]报道了异甾体生物碱抗番茄黄化曲叶病毒(TYLCV)的活性,显示40 μg/mL时3-angeloylzygermine和3-angeloylgermine对TYLCV抑制率分别为74.6%和63.4%,强于阳性对照宁南霉素(51.4%)。
3.8 抗肿瘤作用
转录因子激活蛋白1(AP-1)调节多种蛋白质编码基因,参与许多细胞功能,包括增殖、转化、上皮间质转化和凋亡。研究表明,veratramine型生物碱能对AP-1信号传导途径选择性抑制或直接与AP-1中目标DNA序列的特定位点结合。AP-1下游基因位点Cd14、Mmp9、Il6等在AP-1蛋白被激活时,可受到veratramine型生物碱的抑制;而其他基因位点Fos、Jun和Myc等则受其调节激活,即veratramine型生物碱与AP-1位点的竞争结合,调控着AP-1家族蛋白的表达。而且veratramine型生物碱也能有效抑制SUV(太阳紫外线)诱导的AP-1活化,可在UV诱导的皮肤炎症和致癌中起着重要作用[66]。天目贝母中veratramine型生物碱B18和B19均对HepG2细胞增殖有抑制作用(B18:IC50=1.73 μM;B19:IC50=1.41 μM),且B19也显著抑制Hela细胞增殖(IC50=0.23 μM)[12]。另外环巴胺及其衍生物的肿瘤抑制与NF-κB信号通路、Shh信号通路和Bcl-2的下调等相关,并能通过拮抗Smo受体阻断Hh通路达到抗癌效果。环巴胺及其衍生物CUR0199691能够抑制雌激素受体(ER)的阳性细胞或ER阴性乳腺癌细胞两者之一的增殖揭示了其多靶向性抗癌优势[67]。目前研究发现异甾体类生物碱抗肿瘤的作用机制多与Hedgehog信号通路、caspase-3凋亡、细胞周期和自噬有关[68]。
3.9 其他生物活性
除上述药理活性外,异甾体生物碱还具有抗糖尿病[22]、抑制氧化应激[69]、抗锥虫[70]等生物活性。
4 结语与展望
异甾体生物碱是百合科贝母属和藜芦属植物的主要次生代谢产物,其在抗炎、镇痛、降压、平喘、抗血栓、胆碱酯酶抑制、抗病毒和抗肿瘤等方面具有广泛的应用前景。但多数异甾体生物碱类成分在植物中含量过低、分离难度大制约着异甾体生物碱构效关系的深入开展;还有研究内容大多停留于单一靶点和单一活性,缺乏对其多靶向作用机制的系统研究。为此,笔者建议以下几个研究方向的探索需加以深究:(1)继续对成分不明的同属植物开展化学研究,提升异甾体生物碱结构多样性;探讨同种植物不同部位化学成分及其药用价值,尤其是贝母属药材的研究部位几乎都是鳞茎,花、果、茎、叶等其他部位研究鲜见涉及;基于molecular networking 、LC-MS 、ligand fishing等技术手段进行目标化合物群的精准分离。(2)有效异甾体生物碱(如环巴胺)的简短全合成路线和方法亟需攻克,高含量异甾体生物碱与高活性异甾体生物碱间半合成的转化修饰值得研究。(3)加强异甾体生物碱其他药理活性的筛选,扩大该类成分的应用范围,实行“一物多筛”;对具抗炎、抗肿瘤等多种药理作用的单一成分进行共同作用机制沟通环节的探究,以利发现新颖靶标或未知活性。目前异甾体生物碱的研究仍处于初级阶段,随着新发现的异甾体生物碱不断增多和药理活性的再评价,异甾体生物碱的药用价值定能得以充分开发和利用。
猜你喜欢
中国马铃薯(2022年2期)2022-07-05
中草药(2022年5期)2022-03-03
昆明医科大学学报(2022年1期)2022-02-28
载人航天(2021年5期)2021-11-20
建材发展导向(2021年7期)2021-07-16
中学课程辅导·教学研究(2017年29期)2018-02-26
分析化学(2014年10期)2014-10-24
中国民族民间医药·下半月(2014年4期)2014-09-26
中国民族民间医药·下半月(2014年4期)2014-09-26
时代英语·高三(2014年5期)2014-08-26
