
平潭岛植物群落乔木层碳储量及影响因素研究
2022-02-21 11:04赖敏英肖集泓王春晓邓传远
山东林业科技 2022年6期
赖敏英,肖集泓,李 媚,王春晓,邓传远
(福建农林大学园林学院,福建 福州 350002)
自工业革命以来,人类活动和化石燃料的剧增,导致大气中的二氧化碳等温室气体浓度不断升高,为改善这一状况,人类一方面进行节能减排,另一方面就是植树造林固定大气中的二氧化碳,原因是森林生态系统一方面通过物质和能量循环维持着巨大的碳库,另一方面还吸收约33%由人类活动排放的碳[1-2]。森林植被在光合作用点的过程中吸收二氧化碳,并将其转化为生物量长期地固定在土壤和植被中以达到碳汇的作用[3],其碳贮库由植被(约占42%~50%)、枯死木和枯枝落叶(约占11%~13%)和土壤(约占44%~45%)[4]3个分库组成,这3个库的碳密度和各个林分的面积决定了碳贮[5]。在植被层中,又可以分为乔木层、灌木层和草本层,乔木作为森林植被的主体,森林生态系统中地上部分的碳储量大部分由乔木提供[6-8],仅次于土壤层的碳储量,因此乔木层碳储量值得重视。环境因子、生物因子、土壤因子以及植物的功能性状[9]都可能对植物的碳储量造成影响。计算森林乔木层中的碳储量,分析出森林乔木碳储量与周围环境的相关性,目前已成为森林生态系统碳循环研究的热点问题[10]。
在全球陆地面积中,全球岛陆面积约占1/15,海岛陆地生态系统包括森林、灌木、草本和土壤等,对全球碳循环有重要影响[11]。我国是海岛大国,拥有1万多个大小不同的大陆型岛屿[12],目前对森林生态系统碳储量的研究仍然集中在陆地生态系统,对海岛生态系统碳储量的研究依旧比较薄弱。由于海岛特殊的地理位置、有限的陆地面积,海岛上的植物种类丰富程度较大陆森林生态系统低,群落结构也较为简单,因此在目前的海岛碳储量计算中多运用样地清查结合生物量模型对海岛森林生态系统的碳储量进行计算,研究的海岛包含山东的庙岛群岛[10][39]、舟山群岛[12]、上海崇明岛[13-15]、广东淇澳岛[16]等。也有少数运用遥感对海岛植被的碳储量进行估算[17]。对海岛森林生态系统碳储量进行研究具有可以更好地了解该我国海岛的碳收支情况,也为更加全面地评价海岛生态系统功能提供借鉴意义。平潭岛作为福建第一大岛、我国的第五大岛,以往对平潭岛的研究中,更多的是针对平潭岛海岛典型群落特征、物种多样性以及植被恢复等方面进行研究[18-21],而对平潭岛陆地森林植被的固碳研究较少,且主要针对一种植物的碳储量[17]或是集中植物的碳氮磷化学计量[22]进行研究,对影响该海岛植被碳储量的影响因素的分析仍然比较匮乏。木麻黄和台湾相思作为平潭岛的主要造林树种,对于防风固沙、保护土壤、固氮释氧等各方面具有重要作用,但近几年也面临着更新困难、老化、退化等一系列问题[22]。本研究以木麻黄林、台湾相思林和台湾相思×木麻黄混交林为研究对象,通过实地调查法与生物量相对生长方程相结合的方法,分别计算出3种林分类型的碳生物量,通过乘以含碳系数最终得到3种林分类型的总的碳储量,并运用统计分析方法分析出影响平潭岛乔木层碳储量的主要因子,旨在为当地森林碳汇估算以及促进海岛森林资源可持续经营管理提供基础数据。
1 材料与方法
1.1 研究区域概况
平潭岛(25°15′—25°45′N,119°32′—120°10′E )位于福建省平潭综合实验区,面积占地 324.13 km2,海岸线长408 km,平潭岛地势低平,中部略高,地形以海积平原为主,岛内的最高海拔434.6m,位于君山。平潭岛的气候为南亚热带海洋性季风气候,年平均气温19.0℃~19.9℃,年均降水量900~1200mm,年平均日照约1920小时,夏季风以西南风为主,其余季节风向多为东北风,年平均风速为6.9m/s,平潭岛作为福建省最强风的地区之一,海湾地区全年7级以上大风日数达到125天[23]。平潭岛土壤种类丰富,土壤类型多以砖性红壤和沙壤为主,沙化严重,土壤贫瘠,生境脆弱,研究区域内现有植物中台湾相思(Acacia confusa)纯林、木麻黄(Casuarina equisetifolia)纯林和台湾相思×木麻黄(Acacia confusa×Casuarina equisetifolia)混交林占比较大,伴生有滨柃 (Eurya emarginata)、雀梅藤 (Sageretia thea)、福建胡颓子 (Elaeagnus oldhami)、海桐(Pittosporum tobira) 等灌木和鬼针草 (Bidens pilosa)、 酢浆草 (Oxalis corniculata)、 求米草(Oplismenus undulatifolius)、火炭母(Polygonum chinense)等草本植物。
1.2 研究方法
1.2.1 样地设置
采用植被调查法,根据平潭岛植被分布特征,于2019年9月—2020年9月对平潭岛上的两座内陆山体(瑞岭山和东屿)、一座临海山体(白犬山)和周边3个附属小型海岛(大嵩岛、黄门岛、北香炉屿)进行实地勘察(图1)。本研究选择3种典型次生森林群落作为研究对象,分别为台湾相思林、木麻黄林和台湾相思×木麻黄混交林。台湾相思林和木麻黄林分别设置8个20×20m的样方,台湾相思×木麻黄混交林设置4个20×20m的样方,分别对样地的经纬度、坡度、海拔、坡向等信息进行记录,参照方精云乔木层的测量方法,对群落中所有胸径≥3 cm或树高≥3m的林木进行每木检尺,并记录植物物种名、胸径以及树高等信息,各林分类型具体信息见表1。

表1 样地基本情况Table 1 The basic characteristics of the sampling stations
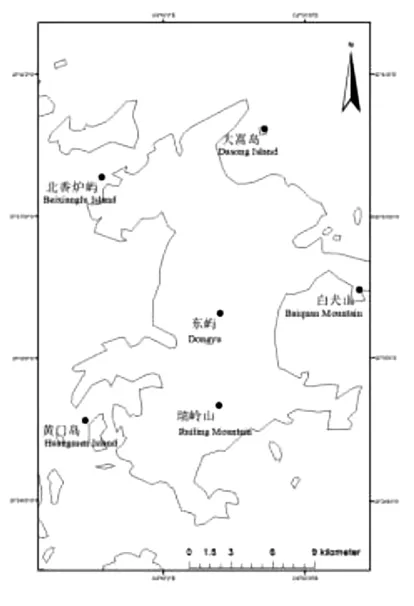
图1 平潭岛乔木层森林乔木层样方分布Figure 1 Locations of the sample plots in the Pingtan Island
1.2.2 土壤样品采集
在每个样地内,分别在样方的4个角以及中心位置设置钻取25 cm深的土样,经过风干、过筛、研磨、去除杂质后,利用四分法将样方内的土壤样品均匀混合成一个土壤样品,对其化学指标进行测定。土壤理化性质按《土壤农业化学分析方法》[24]进行,包含土壤的PH值、有机质、水溶性盐总量、全氮、全磷、全钾、碱解氮、有效磷和速效钾共9项。
1.2.3 乔木层生物量计算
根据测得的乔木数据,选择适合该研究区域的台湾相思、木麻黄的生物量相对生长方程,台湾相思、木麻黄分别参考竹万宽[25]、洪元程[26]建立的生物量相对生长方程(表2)。对3种林分类型的乔木层各个器官(树干、树枝、树叶、树根)的生物量根据公式进行计算,最终将各器官生物量相加得到单位面积内生物量总量(t/hm2)。

表2 木麻黄、台湾相思生物量估算模型Table 2 Biomass estimation model of Casuarina equisetifolia and Acacia confusa
1.2.4 乔木层碳储量计算
本研究中的碳储量仅包含森林植被中乔木层的碳储量,未包含灌木层、草本层和枯枝落叶层以及土壤中的碳储量。生物量乘以含碳系数得到碳储量,台湾相思和木麻黄含碳系数参考《碳汇造林项目方法学》分别按0.485、0.498来计算各个器官的碳储量,各个器官的生物量相加可得到单位面积的生物量,各个单位面积的生物量相加乘以含碳系数可得到各个林分类型乔木层的碳储量。
1.2.5 数据处理
采用Excel 2010软件进行生物量、碳储量计算,采用SPSS 22.0进行相关性分析、单因素(OneWay ANOVA)方差分析和因子分析。
2 结果与分析
2.1 碳储量组成分析
如表3所示,在3种森林类型中以木麻黄林乔木层的碳储量最高,单位面积碳储量达到67.41 t/hm2,其次是台湾相思×木麻黄混交林单位面积碳储量为53.17 t/hm2,高于北长山岛黑松×刺槐混交林(29.60 t/hm2)[10],但也远远低于广西西南部的马尾松×红椎混交林(126.40 t/hm2)[27],台湾相思林乔木层单位面积碳储量为48.91 t/hm2,与张尚炬[28]对沿海台湾相思次生林碳储量(46.54 t/hm2)的测定结果基本一致。木麻黄林碳储量分别比台湾相思林、台湾相思×木麻黄混交林高了37.82%、26.78%。由表3可知,在3种林分类型中,树干的碳储量在总碳储量的占比中都是最大的,树干的碳储量分配为木麻黄林>台湾相思×木麻黄混交林>台湾相思林,尤其是木麻黄林和台湾相思×木麻黄混交林占比分别达到了60.18%和59.07%,说明树干是影响植物碳储量的重要因素;三种林分类型枝、叶的碳储量占比以台湾相思林为最高,分别为18.38%和22.02%;3种林分类型中根的碳储量占比差距较小,均在12.04%~14.26%,其中以木麻黄的根系的碳储量占比最大,这与木麻黄根系深广、抵抗风沙和耐盐碱能力强等特性有关。

表3 不同树种各器官碳储量分配(t/hm2)Table 3 Carbon storage allpcation in different organs in different tree species(t/hm2)
2.2 碳储量的影响因素分析
为找出影响乔木层碳储量的因子,对两个环境因子(坡度、坡向)、3个生物因子(林分密度、平均胸径、平均树高)、8个土壤因子(PH值、水溶性盐含量、有机质、全氮、全钾、碱解氮、有效磷、速效钾)与碳储量进行相关性系数矩阵计算(表4)。结果表明,乔木层碳储量与立地因子、生物因子以及土壤因子具有相关性。在环境因子中,乔木层碳储量主要与坡向具有直接显著正相关性(0.466,p<0.05),说明坡向越向阳、干热程度越大,乔木层碳储量越大;在生物因子中,乔木层碳储量与林分密度(0.543,p<0.05)、平均胸径(0.514,p<0.05)均呈显著正相关关系,与平均树高呈极显著正相关关系(0.617,p<0.01);在土壤因子中,乔木层碳储量与土壤因子无直接显著相关关系。
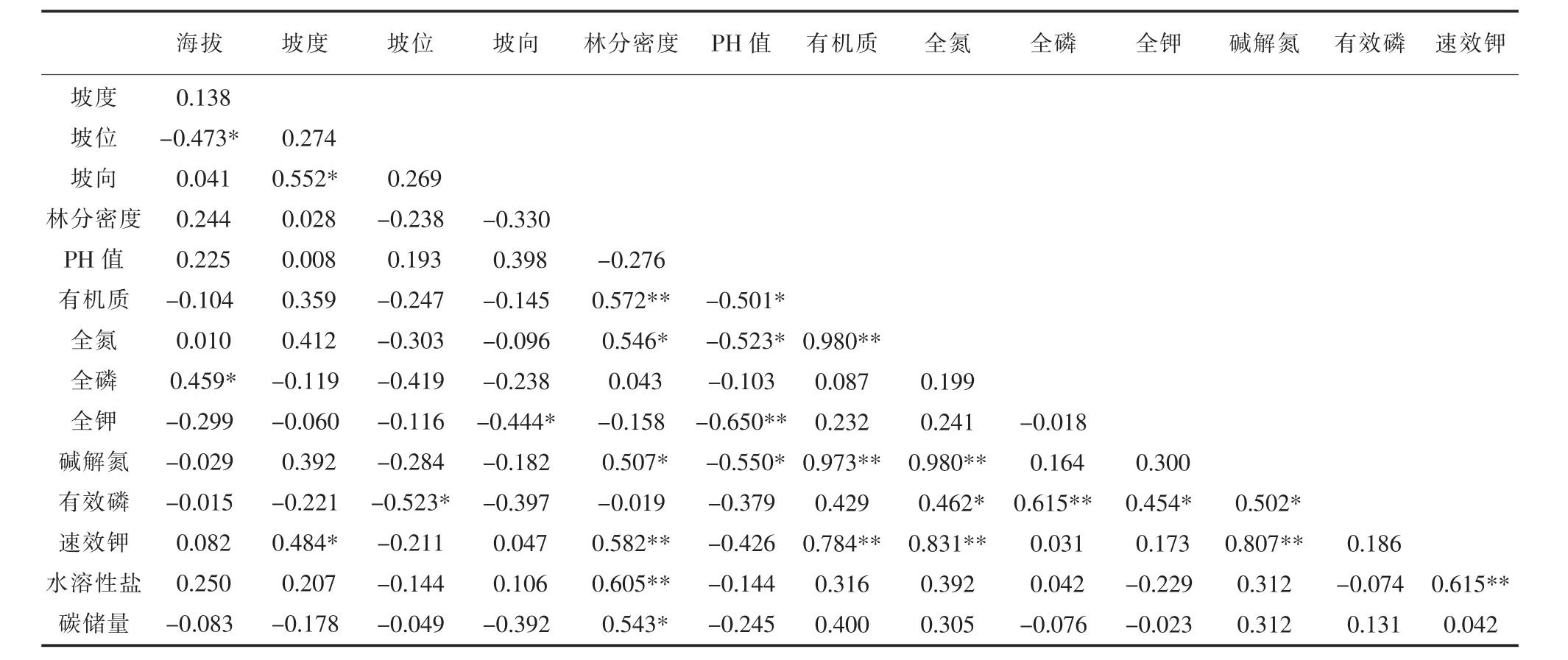
表4 乔木层碳储量与地形因子相关系数矩阵Table 4 Correlation coefficient matrix between tree layer carbon storage and terrain factors
利用SPSS 22.0软件对其进行因子分析,共提取出4个因子(表5),累积贡献率达到84%,说明可以较大程度地说明影响碳储量的影响因素。其中因子1具有较高荷载的是全氮(0.931)、有机质(0.919)、碱解氮(0.901)、速效钾(0.862)、林分密度(0.775)和水溶性盐总量(0.622);因子 2具有较高荷载的是平均胸径(0.944)、平均树高(0.786)和有效磷(0.516);因子 3 具有较高荷载的是全钾(0.930)和 pH(-0.713);因子 4具有较高荷载的坡向(0.843)和坡度(-0.835)。通过多元线性回归表明(表6),各因子的重要程度由高到低依次为因子 2(Beta=0.488),因子 4(Beta=0.462),因子 1(Beta=0.430),因子 3(Beta=-0.241)。 因此在对乔木层碳储量的影响因子中,起主要作用的是生物因子中的平均胸径和平均树高,其次为环境因子(坡度和坡向),再次为土壤土壤因子中各营养元素的含量,而土壤因子中的全钾和PH值对乔木层碳储量的影响程度较小。

表5 乔木层碳储量控制变量主成分分析旋转因子负荷矩阵Table 5 Rotated component matrix derived from a factor analysis of variables controlling arbore carbon storage

表6 乔木层碳储量影响因子得分回归系数Table 6 Regression coefficient of influence factor score of carbon storage in tree layer
虽然在乔木层碳储量与土壤理化性质的相关性分析中,乔木层碳储量与各土壤营养元素不存在直接显著相关性,在对因子分析中土壤因子也不是对乔木层碳储量造成影响的主要因子,但通过对各个森林类型的土壤营养元素进行单因素(OneWayANOVA)方差分析,结果表明(表 7),在 pH值、有机质、全氮、全钾、碱解氮、有效磷、速效钾和水溶性盐总量8项土壤养分指标中,3种林分类型的PH值和水溶性盐总量差异性不显著(p>0.05)外,台湾相思×木麻黄混交林与木麻黄林在有机质、全氮、全钾、碱解氮、有效磷、速效钾6项指标上均存在显著差异 (p<0.05);台湾相思×木麻黄混交林与台湾相思林在全钾和碱解氮两项指标上也存在显著差异(p<0.05)。这种差异是由不同树种类型导致林下植被多样性、凋落物组成、根系的生长发育状况以及分解速率不同引起的[29-30]。土壤的有机质、碱解氮、有效磷和速效钾含量是衡量土壤肥力的重要指标,且指标越大土壤肥力就越高,同时混交林的土壤PH值与木麻黄林和台湾相思林相比都有所减小,土壤含盐量降低,可见,木麻黄与台湾相思混交一方面有利于增加土壤肥力,另一方面对于缓解土壤盐碱化也有重要作用。

表7 3种林型土壤理化性质分析Table 7 Analysis of soil physical and chemical properties of three forest types
3 讨论
本文通过对平潭岛踏勘调研,利用样方调查与生物量相对生长方程相结合的方法,对调查样方的乔木层碳储量进行了研究。在调查的样方中,木麻黄林、台湾相思×木麻黄混交林和台湾相思林中,以木麻黄林碳储量最高,为67.41 t/hm2,高于台湾相思林,介于海南文昌岛东林场木麻黄林[31]的 中 龄 林 (56.00 t/hm2) 与 近 熟 林(99.80 t/hm2)之间,高于福建的一些其他树种乔木层的碳储量,如杉木(50.28 t/hm2)[32]、福建柏 58.70 t/hm2和火力楠 52.66 t/hm2[33]、毛竹林(40.00 t/hm2)[34],也高于福建省森林平均碳密度(48.87 t/hm2)[35],但与世界平均水平(84.00 t/hm2)[36]相比还是有差距。 这与木麻黄作为速生树种,生长速度快,在相同时间较非速生树种积累了更多的生物量以及内抗逆性强、防护效能良好,在风沙地、盐碱地和干旱区生仍然能够长良好有关。
造成植被碳储量差异的影响因子有很多,本研究从环境因子(坡度、坡向)、生物因子(林分密度、平均胸径、平均树高)和土壤因子(PH值、有机质、水溶性盐总量、全氮、全钾、碱解氮、有效磷、速效钾)共3个方面13个因子进行分析。首先在环境因子方面,坡度、坡向等地形条件主要是通过改变生态因子,例如光照、温度、水分、土壤和养分,影响植物的分布和生长发育,进而影响植物碳汇功能的发挥[37]。在沿海地区,风向是影响植物生长的主要干扰因子之一,在林文泉[38]等人对木麻黄立地质量和立地类型的划分的研究中表明,木麻黄在背风向生长较为良好,这是由于坡向对风向和风速影响显著,在背风坡地区,树木生长较迎风坡生长旺盛且树形较迎风坡高大,这也是坡向成为影响乔木层碳储量最明显的立地因子的原因[39],在本研究中,坡向对森林碳储量的影响大于坡度,这一研究结果与许昊、李盈[40-41]等的研究结果接近。在生物因子方面,生物因子能够通过改变林分小气候条件等,进而影响森林生产力及其碳储量[42]。树高和胸径作为计算植被生物量的重要参数,自然与碳储量具有显著的正相关性,因此在本研究中,平均树高和平均胸径是影响乔木层碳储量的主要因子。除树高和胸径以外,林分密度也是影响植被碳储量的生物因子之一。在大多数的研究中,乔木层的碳储量随林分密度的增大而增加[42],在本研究中乔木层的碳储量与林分密度也呈现出显著正相关关系但这都是在林龄较小的情况下,是否林分密度越大乔木层碳储量和生物量越大仍需进一步论证[43]。除了地形因子和生物因子以外,土壤作为陆地生态系统物质循环、水分平衡、凋落物分解等过程的主要载体[44],土壤的养分状况也是影响森林植被碳储量的关键因素,其理化性质对植物的生长发育、更替演化、固碳效益具有重大影响。土壤的化学性质既是响土壤的养分状况的重要因素,也是衡量土壤肥力状况的重要依据[45]。通过分析,土壤化学性质中有效磷是影响森林碳储量的主要因子之一,其次为氮肥、有机质含量、钾肥和土壤含盐量。氮、磷、钾肥作为植物所必须的3大元素,其有效量是易被植物吸收利用的部分[46],植物的生长过程中不可缺少的重要元素。
通过对土壤营养元素的分析发现,台湾相思×木麻黄混交林与木麻黄林营养元素的含量存在显著性差异,与台湾相思林也存在较大差异,台湾相思×木麻黄混交林土壤养分较木麻黄林和台湾相思林有一定的提高,说明混交林对解决土壤结构单一、提升土壤肥力具有具有重要作用[47-48]。
4 结论
虽然在3个林分类型中,木麻黄纯林的碳储量高于台湾相思纯林和台湾相思×木麻黄混交林,更适应在亚热带沿海地区生长,起到防风固沙、良好固碳的理想型树种,但木麻黄林和台湾相思混交并施以相应的肥料(如磷肥),一方面可以改善土壤的物理和化学性质、提升土壤肥力进而减少水土流失、增强对病虫害和自然灾害的抵御能力,另一方面,对于提升台湾相思纯林的固碳能力、生产能力、丰富物种多样性等方面也起着重要作用。
猜你喜欢
建筑与预算(2022年6期)2022-07-08
今日中国·西班牙文版(2021年4期)2021-04-09
农村实用技术(2020年1期)2020-12-16
中学生博览(2020年23期)2020-12-10
绿色科技(2019年13期)2019-08-31
水土保持研究(2019年3期)2019-05-22
天然产物研究与开发(2018年3期)2018-05-07
现代园艺(2018年2期)2018-03-15
现代园艺(2018年14期)2018-01-18
闽南风(2017年12期)2018-01-04
