
瑞香素药动学、毒理学、生物活性的研究进展
2022-02-25 09:38樊思越王文昊罗宁烽张天培高佩佩龙建纲
中成药 2022年2期
樊思越, 王文昊, 罗宁烽, 张天培, 王 珍, 高佩佩, 龙建纲
(西安交通大学,陕西 西安 710049)
瑞香素属于苯并吡喃酮类化合物,又名7,8-二羟基香豆素、祖师麻甲素,是从中药祖师麻中提取得到的一种生物活性单体,同时也是我国自主研发的天然药物,临床上具有镇痛、镇静效果,同时也广泛用于治疗血栓闭塞性脉管炎、冠心病心绞痛、类风湿性关节炎等疾病[1],主要通过抗炎、抗氧化、代谢调节、自噬调节等分子通路发挥作用。本文就瑞香素近年在药动学、毒理学、生物活性方面的研究进展进行综述。
1 理化性质
瑞香素分子式为C9H6O4,化学结构见图1,室温下呈灰白色粉末状,无色无味,为脂溶性药物,碱性条件可发生开环降解,中性或弱酸性条件下较稳定。
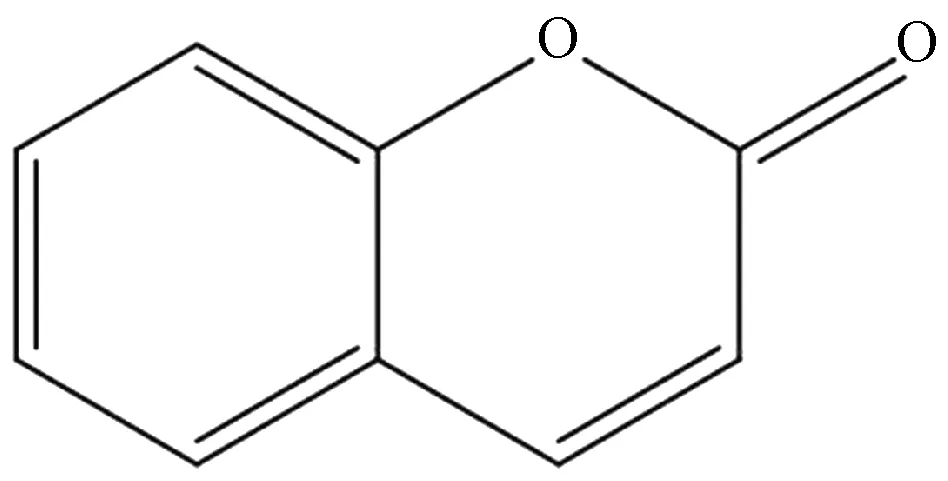
图1 瑞香素化学结构
瑞香素因具有苯并吡喃酮的结构而被归为香豆素类化合物,其母核香豆素的结构如图2所示。由于环上不同位置常含不同取代基,除简单香豆素外,香豆素类化合物还包括呋喃香豆素、吡喃香豆素、其他香豆素,在自然界中分布范围较广,常以游离态或与糖类结合为苷的形式存在于瑞香科、豆科、伞形科等高等植物体内。以往研究及临床应用显示,香豆素类药物具有抑菌、抑肿瘤、抗炎、抗氧化、抗水肿、增强免疫、保肝等生物活性[2-3],其中瑞香素抗炎、抗氧化的生物活性尤为突出。
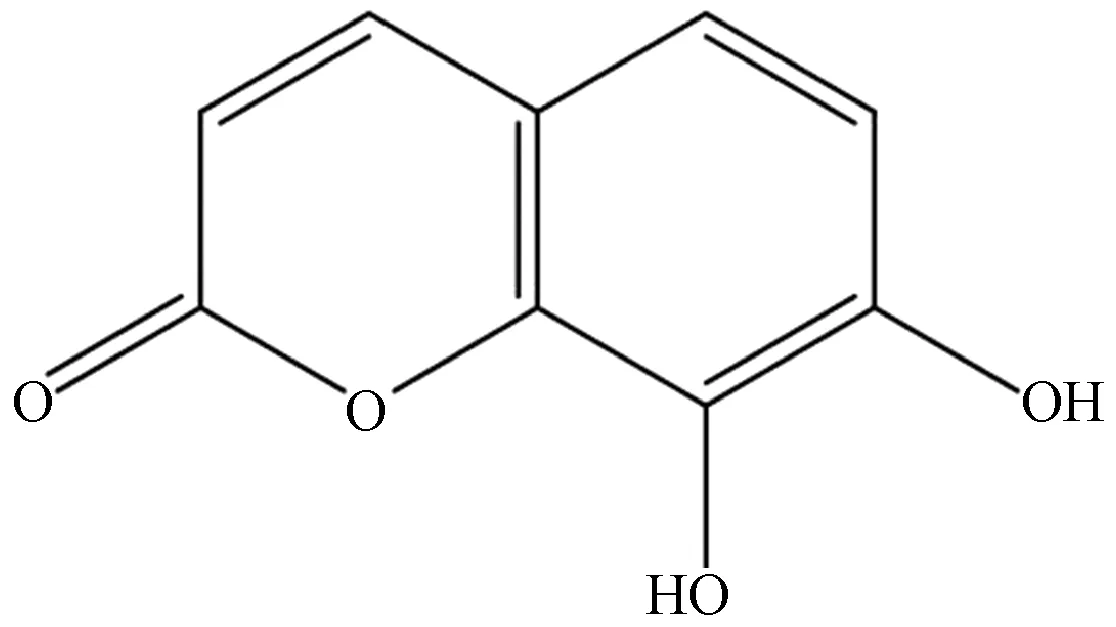
图2 香豆素化学结构
2 药动学
2.1 吸收 曲淑岩等[4]通过大鼠尾静脉给药(80 mg/kg)发现,血药浓度-时间曲线符合二室开放性模型,t1/2=0.043 h,t1/2=0.347 h;家兔经皮给药后,血药浓度-时间曲线符合二室开放性模型,t1/2(ka)=0.462 h,t1/2=0.975 h,t1/2=4.926 h。在中性或酸性条件下瑞香素较为稳定,不易受肠道酶影响,但在弱碱性条件下其内酯结构可能发生开环降解。大鼠在体单向肠灌流模型进行的实验显示,在瑞香素质量浓度10~40 μg/mL范围内,各小肠段的有效渗透系数Peff有上升趋势,但随药物浓度升高无饱和现象,而空肠、结肠Peff有显著性差异 (P<0.05);瑞香素在小肠吸收较好, 而且无特殊的吸收窗[5]。瑞香素在大鼠肠道内可能主要以被动扩散的方式被吸收,同时也可能存在多药耐药相关蛋白2(multidrug resistance-associated protein2, MRP2)、乳腺癌耐药蛋白(breast cancer resistance protein, BCPR)的外排。
2.2 分布 以80 mg/kg剂量对大鼠进行静脉注射给药,15 min后发现瑞香素体内分布广泛,在肝脏、肾脏中浓度最高,肺、脾、血浆、脑中次之,其他器官中也可见少量分布,1 h后不同程度降低,灌胃给药2 h后的分布与静脉给药相似[4]。值得注意的是,瑞香素能穿过血脑屏障,这为中枢神经疾病的防治提供了可能。
2.3 代谢 体外温孵实验证明,全血、肝脏、肾脏都能程度不同地对瑞香素进行代谢,在小鼠体内代谢则主要在肝脏中进行[4,6]。体外代谢特征分析显示,大鼠肝脏中最占优势的代谢途径为磺酸化反应,其次为甲基化和葡糖醛酸化反应,在大鼠肝匀浆上清液中瑞香素7、8位上的羟基易发生上述代谢反应生成相应代谢产物,该过程可能受尿苷二磷酸葡萄糖醛酸转移酶(UDP-ducuronosyltranskrase, UGT)、儿茶酚转甲基酶(catechol-O-methyltransferase, COMT)、磺基转移酶(sulfotransferase, ST)等催化[6];在人类肝脏匀浆上清液中,上述3种代谢途径仍为主要路径,但其中瑞香素清除率最高的途径为甲基化反应,而非磺酸化反应[7]。
2.4 排泄 在静脉注射、灌胃给药2种途径中,瑞香素均主要通过肾脏排泄[4]。大鼠灌胃给药后对其尿液进行检测,约2 h后可发现瑞香素分子的存在,然后其排泄量先增后降,至24~48 h尿液中仅有少量,最终尿液中无法再检测到,其总排泄量不高于最初给药量的6%,其原因可能是该成分转化为其代谢产物[8]。由此说明,瑞香素在大鼠体内消除迅速,24 h内原型药物可基本代谢完全。
3 毒理学
采用最大剂量法(100 mg/kg)对小鼠灌胃给予瑞香素,进行急性毒性试验,观察其皮肤、眼睛、黏膜、呼吸、循环、中枢神经系统,结果连续14 d均未发生中毒迹象,表明该成分对小鼠的口服毒性最大剂量超过100 mg/kg,此质量浓度约为人体有效剂量的175倍[9]。另对50只小鼠腹腔注射瑞香素,测得其LD50为429 mg/kg。
瑞香素溶液对家兔皮肤或粘膜没有刺激作用,皮肤无红斑、浮肿,全身无致敏现象,显微镜下也未发现明显炎症或组织损伤,口腔黏膜形态完好,无其他口腔黏膜刺激反应。由此认为,该成分无致敏性[9]。
在临床应用中,瑞香素也鲜见不良反应发生。对35例应用瑞香素麻醉患者进行身体检查,发现谷丙酶、麝香草酚浊度、尿素氮均在正常范围内,随访21例均未发生不良反应或后遗症。因此,该成分在临床上短期使用是安全的。
4 药理作用
瑞香素除了在心血管疾病方面的临床应用外,对脂肪肝、炎症反应、脑缺血再灌注、脑功能[10]等也具有一定的改善效应,其主要作用机制包括扩张血管、调节炎性因子、抗氧化、调节糖脂代谢、调节细胞自噬、调节线粒体功能等。
4.1 扩张血管 多年来的临床应用证实了瑞香素治疗血栓闭塞性脉管炎、冠心病的可靠性,该成分能扩张冠状动脉,通过舒张主动脉和外周动脉实现收缩压、舒张压降低,血管外周阻力减小[11]。用瑞香素处理实验性2型糖尿病大鼠模型,发现该成分可明显加快血流速度,微血管管径均明显增大[12]。
大部分香豆素类化合物可通过内皮依赖性及非内皮依赖性方式舒张血管,其中又以非内皮依赖性的机制为主。作为香豆素类化合物,瑞香素可能也同样是以剂量依赖性的方式作用于血管平滑肌细胞,通过抑制其膜上钙离子通道,包括电压依赖的钙离子通道(voltage-dependent calcium channels, VDCC)、受体操纵性钙离子通道(receptor-operated calcium channels, ROCC),达到阻止钙离子跨膜进入细胞的目的,并可能阻断钙离子从肌质网中涌出而达到舒张血管的作用[13]。另外,开放钙激活钾离子通道和5-羟色胺受体的竞争拮抗作用也有可能是香豆素类药物发挥血管舒张作用的部分原因[14]。
4.2 免疫调节 瑞香素可在诸多模型中降低白细胞介素-1β(interleukin-1β, IL-1β)、白细胞介素-6(interleukin 6, IL-6)、肿瘤坏死因子-α(tumor necrosis factor alpha, TNF-α)等多种炎症因子表达,涉及丝裂原活化蛋白激酶(mitogen-activated protein kinase, MAPK)、核因子活化B细胞κ轻链增强子(nuclear factorkappa-light-chain-enhancer of activated B cells, NF-κB)等信号通路的调控,从而起到减轻炎症反应的作用。在体外实验中,有学者报道用瑞香素处理TNF-α诱导人永生角质形成细胞,发现该成分能显著减弱该细胞中IL-1β、IL-6、TNF-α、白细胞介素-8(interleukin 8, IL-8)、单核细胞趋化蛋白-1(monocyte chemotactic protein-1, MCP-1)等炎症因子表达上调,表明它可有效减轻TNF-α诱导该模型产生的炎症反应,而该过程可能涉及到NF-κB信号途径[15]。Zhang等[16]研究瑞香素对骨关节炎的作用,发现该成分可抑制IL-1β诱导的兔软骨细胞中IL-6、白细胞介素-12(interleukin 12, IL-12)、基质金属蛋白酶-3(matrix metalloprotein 3, MMP-3)、基质金属蛋白酶-9(matrix metalloprotein 9, MMP-9)、基质金属蛋白酶-13(matrix metalloprotein 13, MMP-13)表达,并能刺激白细胞介素-10(interleukin 10, IL-10)产生,它对MMP的抑制作用部分受到磷脂酰肌醇-3-激酶/丝苏氨酸蛋白激酶(phosphatidyl-inositol 3-kinase/serine-threonine kinase, PI3K/Akt)信号通路、MAPK和NF-κB信号通路抑制性调节。Li等[17]发现,瑞香素对系统性红斑狼疮小鼠也具有炎症抑制作用,可通过上调A20蛋白表达来实现NF-κB通路下调,提高动物存活率,从而起到良好的抗炎效果。
Shen等[18]报道,瑞香素可抑制内毒素诱导的内毒素血症小鼠的促炎因子产生,并能通过抑制脂多糖(lipopolysaccharides, LPS)诱导激活的Janus激酶(janus kinase, JAK)/信号转导子和转录活化子(signal transducer and activator of transcription, STAT)信号通路及活性氧(reactive oxygen species, ROS)产生而表现出抗炎特性。Liu等[19]发现,瑞香素可降低大脑动脉阻塞和再灌注模型小鼠TNF-α、IL-1β、IL-6过表达并抗细胞凋亡,降低Toll样受体4(toll-like receptor 4, TLR4)表达、核因子κB抑制蛋白α(nuclear factor of kappa light polypeptide gene enhancer in B-cells inhibitor alpha, IκB-α)降解及NF-κB的核转位,显示了该成分在模型中良好的神经保护作用及抗炎效果,其潜在分子机制可能涉及对TLR4/NF-κB介导炎性信号传导通路的抑制。
除了对炎性因子的调节外,瑞香素也可以通过其他方式改善实验模型免疫状态。Wang等[20]报道,瑞香素能减轻实验性自身免疫性脑脊髓炎临床症状,显著抑制Th1、Th17细胞反应,抑制DCs活化、成熟、抗原呈递能力;促进胶原诱导关节炎大鼠的T淋巴细胞增殖,并能提高肿瘤杀伤力,促进大鼠免疫功能[21]。
4.3 抗氧化 瑞香素抗氧化效应与丙二醛(malondialdehyde, MDA)、谷胱甘肽(glutathione, GSH)及多种氧化相关酶类如髓过氧化物酶(myeloperoxidase, MPO)、超氧化物歧化酶(superoxide dismutase,SOD)的活性相关。在LPS、D-半乳糖苷诱导的急性肝衰竭过程中,瑞香素可通过抑制MDA形成来降低MPO水平,改善急性肝衰竭导致的SOD、GSH活性下降,缓解氧化应激对肝脏的损害[22]。刘志勇等[23]以瑞香素处理重症急胰腺炎小鼠,发现MDA水平和MPO活性降低。在高葡萄糖诱导的人肾小球系膜细胞中,瑞香素预处理后能降低ROS、MDA水平,提高细胞中SOD活性,进而抑制高葡萄糖诱导的氧化应激[24]。
在氧化应激情况下,瑞香素作用机制可能与NF-E2相关因子2(nuclear erythroid 2-related factor 2, Nrf2)-Kelch样环氧氯丙烷相关蛋白1(kelch-like ECH-associated protein 1, Keap1)系统关系紧密。Lv等[25]研究瑞香素对砷诱导人肺上皮细胞毒性的保护效果及其分子机制,发现该成分以剂量依赖性方式诱促进Nrf2核转位,降低Keap1蛋白表达,Nrf2与抗氧化元件结合,从而上调抗氧化酶表达;诱导AMP依赖的蛋白激酶(adenosine 5′-monophosphate (AMP)-activated protein kinase, AMPK)活化,c-Jun 氨基端激酶(c-Jun N-terminal kinase, JNK)和胞外信号调节激酶(extracellular signal-regulated kinase, ERK)磷酸化,并能阻碍砷诱导的细胞活力下降和ROS产生,推测其对砷作用产生的细胞毒性和细胞凋亡可能与Nrf2依赖的抗氧化反应的诱导以及抗凋亡因子(B-cell lymphoma-2, Bcl-2)在人肺上皮细胞中的稳定有关。Zhi等[26]发现,瑞香素能通过诱导Nrf2的核易位来诱导血红素加氧酶-1(Heme oxygenase, HO-1)的表达和活性,减轻海马神经元缺血再灌注模型中的氧化应激及细胞损伤。
4.4 调节糖脂代谢 Liu等[27]在油酸诱导的HepG2细胞模型中发现,瑞香素干预后可降低固醇调节元件结合蛋白-1(sterol regulatory element-binding protein-1, SREBP-1)、马铃薯糖蛋白样磷脂酶结构域蛋白(patatin-like phospholipase domain-containing protein 3,PNPLA3)的蛋白和mRNA表达,上调过氧化物酶体增殖物激活受体α(peroxisome proliferator activated receptor-α,PPARα)基因、AMPK磷酸化水平,从而抑制脂质在肝细胞中的积累;增强磷脂酰肌醇激酶PI3K蛋白表达,促进Akt磷酸化,从而促进肝细胞对葡萄糖的吸收和利用。在糖尿病的研究中,PPARs一直被认为是治疗靶点,但已有靶向药物具有肝毒性、体质量增加等严重不良反应,目前开发PPARs双重激动剂,旨在改善代谢同时能尽量减小副作用。有研究表明,瑞香素质量浓度在40、80 μmol/L下可双重激活PPAR-α/γ, 80 μmol/L下可使PPAR-α/β/γ表达得到激活[28-29]。此外,有学者发现瑞香素对链脲佐菌素(streptozotocin, STZ)诱导的大鼠胰岛素瘤胰腺β细胞损伤具有抑制脂质过氧化的效果,并可改善该细胞胰岛素分泌障碍,显示出瑞香素具有抵抗糖尿病的潜力[30]。由此表明,瑞香素在调节糖脂代谢方面有重要临床应用前景。
4.5 自噬调节 Lv等[22]构建急性肝衰竭模型,发现由LPS及D-半乳糖苷诱导的小鼠急性肝衰竭会导致肝脏自噬因子Beclin-1、Agt16、Agt12、Agt7、Agt5、Agt3蛋白表达下降,并会降低LC3Ⅱ与LC3Ⅰ的比率,而瑞香素干预则能明显地抑制上述自噬因子表达的下调,从而起到保护肝脏的作用。Zhang等[31]报道,瑞香素可促进金黄色葡萄球菌处理的巨噬细胞中LC3-Ⅰ向LC3-Ⅱ的转化和LC3颗粒的形成,同时瑞香素抑制雷帕霉素靶蛋白(mammalian target of rapamycin, mTOR)及下游信号通路的激活,抑制转录因子EB(transcription factor EB,TFEB),从而触发溶酶体基因的表达。
除此之外,瑞香素也可在自噬激活模型中发挥抑制作用。Deng等[32]以TNF-α诱导胶原蛋白诱导性关节炎大鼠的成纤维样滑膜细胞,发现瑞香素可减弱TNF-α诱导所致Atg5、Beclin-1、LC3蛋白表达上调,并通过抑制Akt和mTOR活化、下调PKB信号通路激活,从而抑制关节炎大鼠成纤维样滑膜细胞自噬,减少细胞中受TNF-α刺激增加自噬体。
4.6 线粒体相关调节 瑞香素通过促进JNK、ERK的磷酸化激活来使得Nrf2与Keap1之间实现有效分离,然后Nrf2蛋白进一步启动下游启动子转录,从而诱导包含谷氨酸-半胱氨酸连接酶催化亚基(glutamate-cysteine ligase catalytic subunit, GCLC)、谷氨酸-半胱氨酸连接酶修饰亚基(glutamate-cysteine ligase modifier subunit, GCLM)、HO-1在内的一系列抗氧化酶表达,有效改善线粒体功能障碍,表现为抑制ROS对线粒体的氧化损害,从而抑制炎症应答及细胞凋亡[33]。由此表明,瑞香素可能有助于线粒体损伤相关疾病的防治。
5 展望
目前,瑞香素已作为临床治疗脉管炎、冠心病等疾病的药物,可通过免疫调节、糖脂代谢调节、自噬调节等多种分子通路发挥生物学活性。进一步探索该成分作用机制对相关疾病防治具有重要意义。例如,阿尔兹海默症不仅是中枢神经病理变化系统疾病,其早期还会伴随中枢和外周神经系统的慢性炎症、氧化应激,表现出线粒体功能障碍、代谢紊乱等病理症状,而瑞香素的多重药理效应和分子通路有望为该疾病防治提供新的研究思路及解决方案。
猜你喜欢
现代园艺(2022年12期)2022-11-19
中国药业(2022年17期)2022-09-08
保健医苑(2022年4期)2022-05-05
保鲜与加工(2021年12期)2022-01-07
食品安全导刊(2021年20期)2021-08-30
当代水产(2021年6期)2021-08-13
汽车维修与保养(2021年8期)2021-02-16
建材发展导向(2019年10期)2019-08-24
时尚北京(2016年12期)2017-01-16
中国新技术新产品(2015年13期)2015-12-26
