
海洋来源担子菌产葡萄糖氧化酶发酵条件优化
2022-03-09 06:09张庆芳王倩倩迟雪梅陈立功迟乃玉刘春莹
微生物学杂志 2022年5期
张庆芳, 王倩倩, 迟雪梅, 于 爽, 陈立功, 迟乃玉, 刘春莹*
(1.大连大学 生命科学与技术学院,辽宁 大连 116622;2.辽宁省海洋微生物工程技术研究中心,辽宁 大连 116622)
葡萄糖氧化酶(glucose oxidase, EC l.1.3.4, GOD)在分子氧存在下能氧化葡萄糖(glucose, Glu)生成D-葡萄糖酸内酯,同时消耗氧生成过氧化氢,其催化过程或产物有去除葡萄糖、杀菌和脱氧作用,在畜牧养殖、水产、医疗及化工等领域应用广泛[1-2]。GOD广泛分布于自然界生物体内,已研究的GOD主要来源于霉菌和昆虫[3-6],细菌和酵母菌来源的GOD较少[7-12]。目前,随着科学研究的进展,GOD酶活性检测方法也在逐步创新和优化,主要包括电化学法、靛红褪色法和分光光度法等进行GOD酶活性检测[13-15]。电化学法通过测定酶促反应中的电压或电流的变化,推算酶活性[16-19];靛红褪色法利用催化生成的H2O2使靛蓝胭脂红褪色计算酶活性[20-23]。分光光度法是目前使用最多的GOD酶活性检测方法,利用GOD-过氧化物酶-邻联茴香胺偶联反应,测定反应后因物质变化引起的吸光度的改变,进而推算出GOD的酶活性[24-25]。该方法灵敏度高,试剂用量较少,但在本研究检测过程中,发现该方法检测GOD酶活性不稳定,有时甚至测不出酶活性[21,26-27]。本研究对传统分光光度法进行改良,避免邻联茴香胺溶液中的乙醇破坏蛋白质中原有的氢键,影响蛋白质活性。目前,GOD研究主要存在两大问题,一是研究对象多来自陆地环境,例如霉菌和昆虫等,属于中高温酶,而对低温GOD的研究较少[28-29];二是对微生物来源的GOD的研究主要集中在霉菌和细菌,而以曲霉研究最多,对酵母菌来源的GOD研究较少[30-31],尤其是海洋担子菌属来源的GOD的研究更少,已报道的只有刘春莹等[32]从海洋中筛选到1株担子菌。因此,研究海洋担子菌来源的低温GOD,对GOD资源库的扩充具有重要意义。本研究使用的菌株来自渤海海域海泥,是通过前期筛选得到的1株高产低温GOD菌株,经过形态学及分子生物学鉴定确定其属担子菌属(Basidioascussp.),命名为Basidioascussp.LG-31,在前期实验的基础上,对其进行发酵条件优化。为海洋来源的低温GOD研究及低温GOD的工业化生产及应用提供参考。
1 材料与方法
1.1 材料
1.1.1 菌株来源 菌株来自辽宁省海洋微生物工程中心实验室,分离自辽宁大连渤海海域(123°371′E,39°697′N),深度为25~30 m的海泥,命名为Basidioascussp.LG-31。
1.1.2 培养基 发酵/种子培养基(g/L):葡萄糖60,蛋白胨3,KH2PO40.2,MgSO4·7H2O 0.7,KCl 0.5,NaNO34,pH 7.0。
1.1.3 主要试剂与仪器设备 邻联茴香胺溶液、辣根过氧化物酶溶液、5%的葡萄糖溶液(自配);D(+)-麦芽糖(BR)、发酵用麸皮(BR)、发酵用秸秆粉(BR)、玉米浆膏(BR)(北京奥博星生物科技有限公司);D-甘露糖(Biotech Grade)、L-谷氨酸(BC Grade)、M-苯丙氨酸(AR)、N-乙酰-L-苯丙氨酸(AR)、无水肌酸(AR)(生工生物科技(上海)有限公司);葡聚糖(AR)(中国医药(集团)上海化学试剂公司);微晶纤维素(Tech Grade)、柠檬酸三铵(USP Grade)(BBI有限公司);KNO3(AR)(天津市科米欧化学试剂有限公司);NH4NO3(AR)(天津市双船化学试剂厂);NH4Cl(AR)(国药集团化学试剂有限公司);(NH4)2HPO4(AR)、(NH4)2SO4(AR)、乙酸铵(AR)(天津市大茂化学试剂厂);氨水(AR(25%))(天津市凯信化学工业有限公司)。低温恒温培养箱(LTI-700,上海艾朗实验设备有限公司);恒温摇床(CRY-2112,上海艾朗实验设备有限公司);酶标仪(Multiskan GO,赛默飞世尔科技(芬兰)有限公司);立式压力蒸汽灭菌器(YXQ-LS-100SII,上海博讯工贸有限公司);微量高速冷冻离心机(Biofuge fresco,赛默飞世尔科技(中国)有限公司);高速冷冻离心机(CR21N,日立(中国)有限公司)。
1.2 方法
1.2.1 酶活性测定 对传统分光光度法进行改良,避免了邻联茴香胺溶液中的乙醇破坏蛋白质中原有的氢键,影响蛋白质活性。①对照组:将邻联茴香胺溶液150 μL和辣根过氧化物酶溶液10 μL加入96孔板中,并于25 ℃保温10 min,然后加入150 μL 5%葡萄糖溶液和30 μL粗酶液,立刻测OD460,3组平行实验。②实验组:将邻联茴香胺溶液150 μL和辣根过氧化物酶溶液10 μL加入96孔板中,25 ℃保温10 min;将150 μL粗酶液和750 μL 5%葡萄糖溶液混合均匀后于30 ℃水浴保温5 min,取180 μL保温好的粗酶液与葡萄糖的混合溶液加入到邻联茴香胺-辣根过氧化物酶混合溶液中,振荡反应10 s,测OD460,3组平行实验。酶活性单位(U)定义:在上述条件下,催化产生l μmol N-乙酰-D-氨基葡萄糖所需的酶量。酶活性计算公式:Y=[(A-A0)+C]/K×N/T/M。式中,Y:葡萄糖氧化酶活性(U/mL);A:实验组的吸光度;A0:对照组的吸光度;K:标准曲线的斜率;C:标准曲线的截距;N:酶液的总稀释倍数;T:反应时间(min);M:酶液体积(mL)。
1.2.2 生长曲线和酶活性曲线测定方法 挑取菌株Basidioascussp. LG-31单菌落置于5 mL种子培养基中,25 ℃、200 r/min培养至OD600为0.55~0.60,作为测定生长曲线及酶活性曲线的种子液。以1%接种量接种菌株Basidioascussp.LG-31种子液于100 mL发酵培养基中,25 ℃、200 r/min培养,每4 h取菌液测OD600吸光度,3组平行实验,取平均值绘制生长曲线。以1%接种量接种菌株Basidioascussp.LG-31种子液于100 mL发酵培养基中,25 ℃、200 r/min培养,每4 h取发酵液4 000 r/min离心,取上清于OD460处检测酶活性,3组平行实验,取平均值绘制酶活性曲线。
1.2.3 单因素优化方法 ①碳源种类优化:选取葡萄糖、麦芽糖、蔗糖、可溶性淀粉、乳糖、甘油、发酵用麸皮、发酵用秸秆粉、D-甘露糖、葡聚糖、微晶纤维素、几丁质做为碳源。添加量(质量分数,下同)为碳源6%,蛋白胨0.3%,NaNO30.4%,KH2PO40.02%,MgSO4·7H2O 0.07%,KCl 0.05%,发酵温度25 ℃,初始pH 7.0,装液量100 mL/250 mL,接种量1%(体积分数,下同),200 r/min培养28 h,于OD460处检测酶活性。②蔗糖添加量优化:蔗糖添加量分别为2%、4%、6%、8%、10%、12%、14%、16%,蛋白胨0.3%,NaNO30.4%,KH2PO40.02%,MgSO4·7H2O 0.07%,KCl 0.05%,发酵温度25 ℃,初始pH 7.0,装液量100 mL/250 mL,接种量1%,200 r/min培养28 h,于OD460处检测酶活性。③有机氮源种类优化:有机氮源分别为胰蛋白胨、蛋白胨、牛肉膏、酵母粉、酵母浸粉、酵母膏、玉米浆膏、尿素、尿酸、乙酰胺、干酪素、L-谷氨酸、L-苯丙氨酸、N-乙酰-L-苯丙氨酸、无水肌酸,其添加量为0.3%,蔗糖4%,NaNO30.4%,KH2PO40.02%,MgSO4·7H2O 0.07%,KCl 0.05%,发酵温度25 ℃,初始pH 7.0,装液量100 mL/250 mL,接种量1%,200 r/min培养28 h,于OD460处检测酶活性。④酵母粉添加量优化:酵母粉添加量分别为0.1%、0.2%、0.3%、0.4%、0.5%、0.6%、0.7%、0.8%、0.9%、1.0%,蔗糖4%,NaNO30.4%,KH2PO40.02%,MgSO4·7H2O 0.07%,KCl 0.05%,发酵温度25 ℃,初始pH 7.0,装液量100 mL/250 mL,接种量1%,200 r/min培养28 h,于OD460处检测酶活性。⑤无机氮源种类优化:无机氮源分别为NaNO3、KNO3、NH4NO3、NH4Cl、(NH4)2SO4、氨水、(NH4)2HPO4、柠檬酸三铵、柠檬酸氢二铵、乙酸铵,其添加量0.4%,蔗糖4%,酵母粉0.3%,KH2PO40.02%,MgSO4·7H2O 0.07%,KCl 0.05%,发酵温度25 ℃,初始pH 7.0,装液量100 mL/250 mL,接种量1%,200 r/min培养28 h,于OD460处检测酶活性。⑥NaNO3添加量优化:NaNO3添加量分别为0.1%、0.2%、0.3%、0.4%、0.5%、0.6%、0.7%、0.8%、0.9%、1.0%,蔗糖4%,酵母粉0.3%,KH2PO40.02%,MgSO4·7H2O 0.07%,KCl 0.05%,发酵温度25 ℃,初始pH 7.0,装液量100 mL/250 mL,接种量1%,200 r/min培养28 h,于OD460处检测酶活性。⑦KH2PO4添加量优化:KH2PO4添加量分别为0、0.02%、0.04%、0.06%、0.08%、0.10%、0.12%、0.14%、0.16%、0.18%、0.20%,蔗糖4%,酵母粉0.3%,NaNO30.4%,MgSO4·7H2O 0.07%,KCl 0.05%,发酵温度25 ℃,初始pH 7.0,装液量100 mL/250 mL,接种量1%,200 r/min培养28 h,于OD460处检测酶活性。⑧KCl添加量优化:KCl添加量分别为0、0.05%、0.10%、0.15%、0.20%、0.25%、0.30%、0.35%、0.40%,蔗糖4%,酵母粉0.3%,NaNO30.4%,KH2PO40.02%,MgSO4·7H2O 0.07%,发酵温度25 ℃,初始pH 7.0,装液量100 mL/250 mL,接种量1%,200 r/min培养28 h,于OD460处检测酶活性。⑨MgSO4·7H2O添加量优化:MgSO4·7H2O添加量分别为0、0.02%、0.04%、0.06%、0.08%、0.10%、0.12%、0.14%,蔗糖4%,酵母粉0.3%,NaNO30.4%,KH2PO40.02%,KCl 0.1%,发酵温度25 ℃,初始pH 7.0,装液量100 mL/250 mL,接种量1%,200 r/min培养28 h,于OD460处检测酶活性。⑩发酵初始pH优化:蔗糖4%,酵母粉0.3%,NaNO30.4%,KH2PO40.02%,KCl 0.1%,MgSO4·7H2O 0.1%,初始pH 3~11,发酵温度25 ℃,装液量100 mL/250 mL,接种量1%,200 r/min培养28 h,于OD460处检测酶活性。接种量优化:蔗糖4%,酵母粉0.3%,NaNO30.4%,KH2PO40.02%,KCl 0.1%,MgSO4·7H2O 0.1%,初始pH 7.0,发酵温度25 ℃,装液量100 mL/250 mL,接种量分别为0.5%、1.0%、1.5%、2.0%、2.5%、3.0%、3.5%、4.0%、4.5%、5.0%、5.5%、6.0%,200 r/min培养28 h,于OD460处检测酶活性。装液量优化:蔗糖4%,酵母粉0.3%,NaNO30.4%,KH2PO40.02%,KCl 0.1%,MgSO4·7H2O 0.1%,初始pH 7.0,发酵温度25 ℃,装液量分别为25 mL/250 mL、50 mL/250 mL、75 mL/250 mL、100 mL/250 mL、125 mL/250 mL、150 mL/250 mL、175 mL/250 mL、200 mL/250 mL,接种量2%,200 r/min培养28 h,于OD460处检测酶活性。发酵转速优化:蔗糖4%,酵母粉0.3%,NaNO30.4%,KH2PO40.02%,KCl 0.1%,MgSO4·7H2O 0.1%,初始pH 7.0,发酵温度25 ℃,装液量125 mL/250 mL,接种量2%,分别于80、100、120、140、160、180、200、220 r/min培养28 h,于OD460处检测酶活性。发酵温度优化:蔗糖4%,酵母粉0.3%,NaNO30.4%,KH2PO40.02%,KCl 0.1%,MgSO4·7H2O 0.1%,初始pH 7.0,发酵温度分别为5、10、15、20、25、30、35、40、45、50 ℃,装液量125 mL/250 mL,接种量2%,200 r/min培养28 h,于OD460处检测酶活性,所有实验均设3组平行。
2 结果与分析
2.1 生长曲线与酶活性曲线
如图1标准曲线所示,OD460(y)与H2O2(x)含量之间的关系为y=0.005 2x+0.013 7,R2=0.997 4,大于0.99,表明该标准曲线可用。

图1 酶活性标准曲线图Fig.1 Standard curve of enzyme activity
菌株Basidioascussp.LG-31在25 ℃,200 r/min培养条件下,生长曲线和酶活性曲线如图2所示。由生长曲线可知,菌株Basidioascussp. LG-31在0~16 h处于延滞期,生长繁殖缓慢,产酶量少;16 h后菌株生长进入对数期,菌株生长繁殖加剧,发酵液菌体浓度迅速增加,这一阶段GOD酶活性也快速升高,28 h时酶活性达到最高,为15.28 U/mL,而后酶活性迅速降低;约48 h时菌株生长进入稳定期,此阶段菌株代谢活动减慢,增殖速度随之降低,发酵液菌体浓度相对稳定,菌株产酶很少,基本无GOD酶活性;菌株生长进入56 h后,缓慢进入衰亡期,此阶段菌株增殖速度小于死亡速度,可能因为发酵进入此时期时,发酵液内营养物质消耗过多,不能满足大量菌体生长繁殖需要。确定该菌株产酶最适培养时间为28 h。
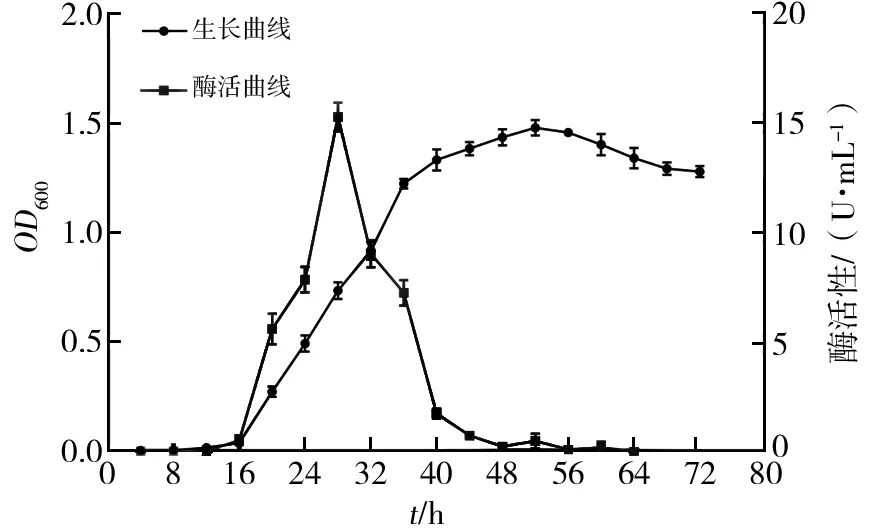
图2 菌株Basidioascuss sp. LG-31生长曲线与酶活性曲线Fig.2 Growth and enzyme activity curves of strain Basidioascus sp. LG-31 strain
2.2 菌株Basidioascus sp.LG-31产GOD发酵条件的单因素优化
2.2.1 不同碳源对菌株Basidioascussp.LG-31产酶的影响 由图3可知,蔗糖是该菌株生长产酶的最佳碳源,可溶性淀粉、葡聚糖对菌株产酶有较好的诱导作用,葡萄糖、乳糖、甘油、发酵用麸皮、发酵用秸秆粉及D-甘露糖有一定的诱导产酶能力。麦芽糖、微晶纤维素和几丁质不能诱导菌株产酶。

图3 不同碳源对菌株Basidioascus sp.LG-31酶活性的影响Fig.3 Effects of different carbon sources on enzyme production by Basidioascus sp. LG-31 strain
2.2.2 蔗糖添加量对菌株Basidioascussp. LG-31酶活性的影响 由图4可知,当蔗糖添加量为2%时,菌株酶活性最高,随着蔗糖添加量的增加,菌株酶活性降低。因此,确定最佳蔗糖添加量为4%。
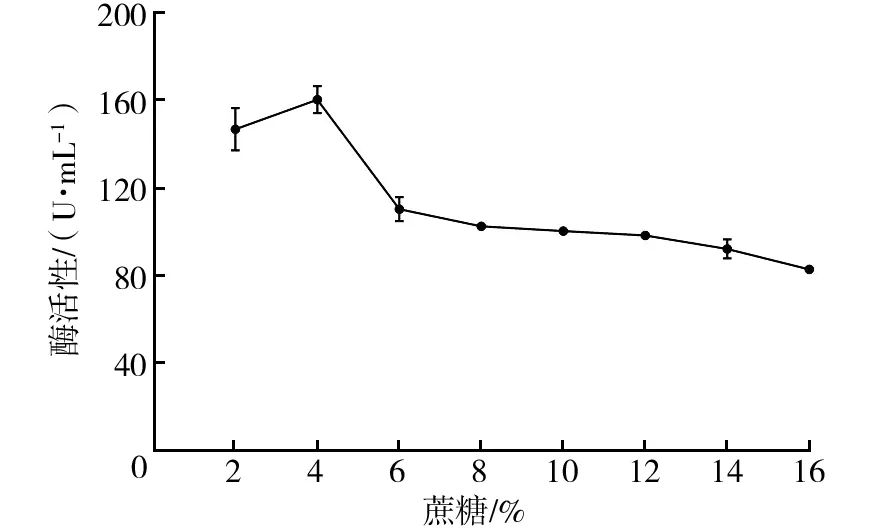
图4 蔗糖添加量对菌株Basidioascus sp. LG-31酶活性的影响Fig.4 Effect of sucrose addition on enzyme production by Basidioascus sp. LG-31 strain
2.2.3 不同有机氮源对菌株Basidioascussp. LG-31酶活性的影响 由图5可知,酵母粉是该菌株生长产酶的最佳有机氮源,与对照组相比,胰蛋白胨、乙酰胺、L-苯丙氨酸、无水肌酸对菌株酶活性均有促进作用;其余有机碳源对菌株酶活性有抑制作用,尿素和L-谷氨酸抑制效果最强。

图5 不同有机氮源对菌株Basidioascus sp.LG-31酶活性的影响Fig.5 Effects of different organic nitrogen sources on enzyme production by Basidioascus sp. LG-31 strain
2.2.4 酵母粉添加量对菌株Basidioascussp. LG-31酶活性的影响 由图6可知,在一定浓度范围内,酵母粉添加量与葡萄糖氧化酶活性呈正比例关系;当酵母粉添加量为0.3%时,菌株产葡萄糖氧化酶的酶活性达到最高,此后继续增大酵母粉添加量,酶活性下降。当酵母粉添加量在0.3%~0.6%时,对菌株酶活性影响不大。因此,确定酵母粉最佳添加量为0.3%。

图6 酵母粉添加量对菌株Basidioascus sp. LG-31酶活性的影响Fig.6 The effect of the added amount of yeast powder on the enzyme production of Basidioascus sp. LG-31 strain


图7 不同无机氮源对Basidioascus sp.LG-31菌株酶活性的影响Fig.7 Effects of different inorganic nitrogen sources on enzyme production by Basidioascus sp. LG-31 strain
2.2.6 NaNO3添加量对菌株Basidioascussp. LG-31酶活性的影响 由图8可知,在一定浓度范围内,随着NaNO3添加量的增加,GOD酶活性升高,当NaNO3添加量为0.4%时,酶活性最高,继续增大NaNO3添加量,酶活性下降。因此,确定最佳NaNO3添加量为0.4%。

图8 NaNO3添加量对菌株Basidioascus sp.LG-31产酶的影响Fig.8 Effect of NaNO3 addition on enzyme production by Basidioascus sp. LG-31 strain
2.2.7 无机盐添加量对菌株Basidioascussp.LG-31酶活性的影响 ① KH2PO4添加量优化:由图9可知,低浓度KH2PO4对菌株产酶有促进作用,当KH2PO4添加量为0.02%时,菌株酶活性最高;随着KH2PO4添加量的增加,酶活性下降。因此,确定最佳KH2PO4添加量为0.02%。② KCl添加量优化:由图10可知,KCl添加量为0.1%时,菌株酶活性最高,随着KCl添加量的增加,酶活性缓慢下降,高浓度的KCl对菌株产酶抑制效果较强。最佳KCl添加量为0.10%。③ MgSO4·7H2O添加量优化:由图11可知,在一定浓度范围内,随着MgSO4·7H2O添加量的增加,葡萄糖氧化酶活性升高,当MgSO4·7H2O添加量为0.10%时,酶活性最高,继续增大MgSO4·7H2O添加量,酶活性下降。因此,确定最佳MgSO4·7H2O添加量为0.10%。
2.2.8 初始发酵pH对菌株Basidioascussp. LG-31酶活性的影响 如图12所示,菌株在发酵培养基初始pH越接近中性时酶活性越高,偏酸或偏碱都会影响菌株酶活性,当pH为7.0时产酶活性最高。因此,确定最佳发酵培养基初始pH为7.0。
2.2.9 接种量对菌株Basidioascussp. LG-31酶活性的影响 如图13所示,接种量为2%时,GOD酶活性最高,当接种量超过2%,GOD酶活性随着接种量的增大急剧降低,可能是因为接种量浓度过大,培养基中菌体生长产酶所需的氧气消耗加剧,不能满足其正常的生长产酶需要。因此,确定最适接种量为2%。
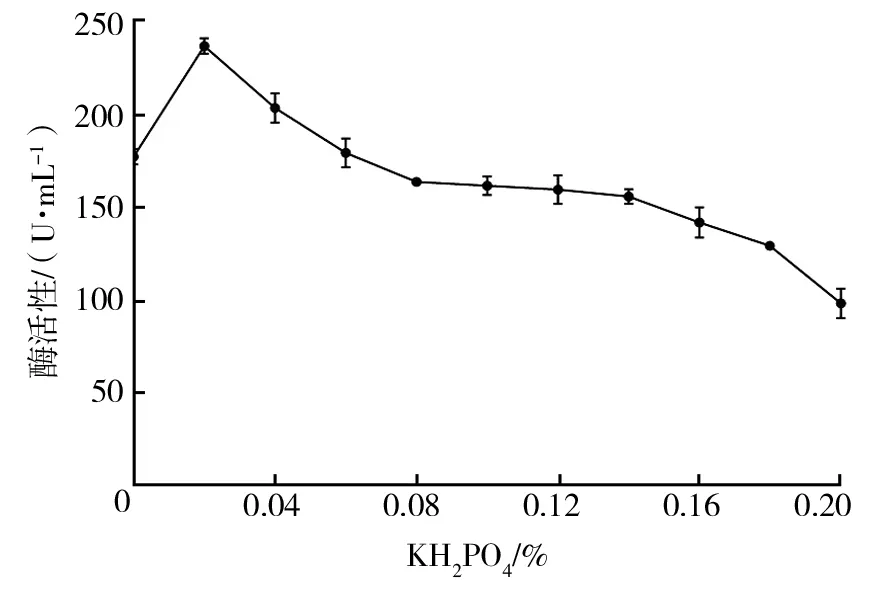
图9 KH2PO4添加量对菌株Basidioascus sp.LG-31酶活性的影响Fig.9 Effects of KH2PO4 addition on enzyme production by Basidioascus sp. LG-31 strain
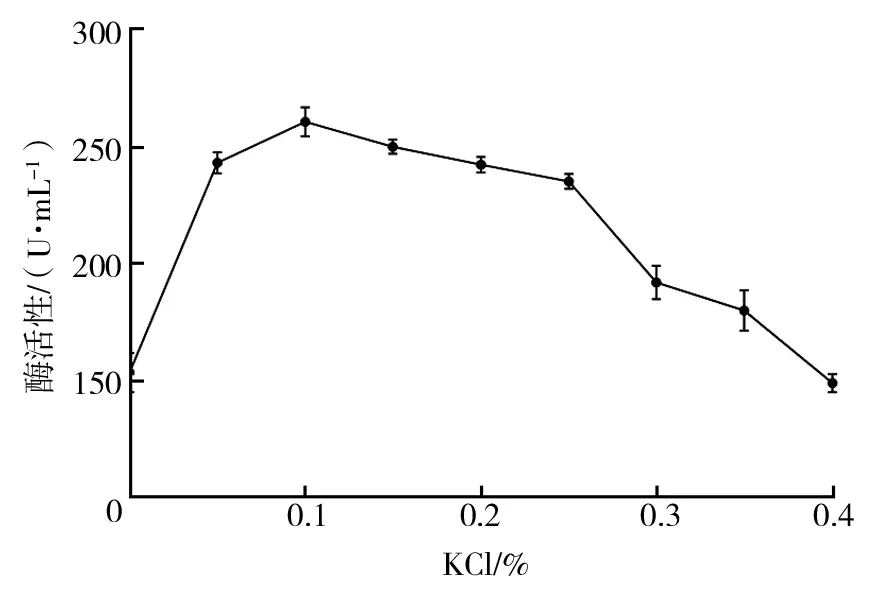
图10 KCl添加量对菌株Basidioascus sp.LG-31酶活性的影响Fig.10 Effects of KCl addition on enzyme production by Basidioascus sp. LG-31 strain

图11 MgSO4·7H2O添加量对菌株Basidioascuss sp. LG-31酶活性的影响Fig.11 Effects of MgSO4·7H2O addition on enzyme production by Basidioascus sp. LG-31 strain

图12 初始发酵pH对菌株Basidioascus sp. LG-31酶活性的影响Fig.12 The effect of initial fermentation pH on enzyme production of Basidioascus sp. LG-31 strain

图13 接种量对菌株Basidioascus sp. LG-31酶活性的影响Fig.13 Influence of inoculation amount on enzyme production of Basidioascus sp. LG-31 strain
2.2.10 装液量对菌株Basidioascussp. LG-31酶活性的影响 由图14可知,当装液量为125 mL/250 mL时,菌株酶活性最高,增加或减少装液量都会使GOD酶活性降低。因此,确定最适装液量为125 mL/250 mL。
2.2.11 发酵转速对菌株Basidioascussp. LG-31酶活性的影响 由图15可知,转速较低时,菌株酶活性随着转速的增加而增高,当转速达到200 r/min时,酶活性达到最高值。因此,确定最适转速为200 r/min。
2.2.12 发酵温度对菌株Basidioascussp. LG-31酶活性的影响 由图16可知,温度过高或过低,都会影响菌株产GOD活性,当发酵温度为15~30 ℃时,菌株酶活性较高,当发酵温度为20 ℃时,菌株GOD酶活性最高,这可能是因为菌株来源于海洋。因此,确定最适发酵温度为20 ℃。

图14 装液量对菌株Basidioascus sp. LG-31酶活性的影响Fig.14 The effect of liquid volume on enzyme production of Basidioascus sp. LG-31 strain
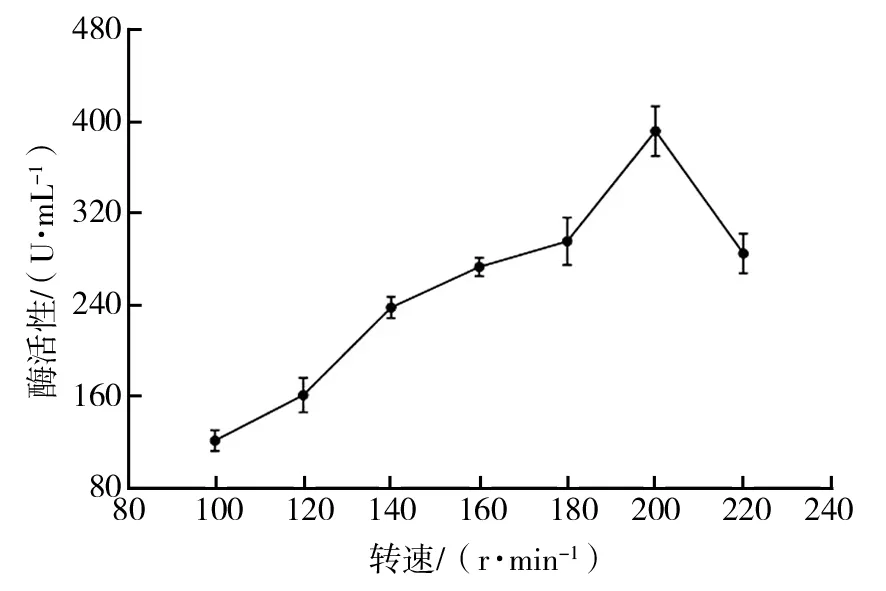
图15 发酵转速对菌株Basidioascus sp. LG-31酶活性的影响Fig.15 The effect of fermentation speed on enzyme production of Basidioascus sp. LG-31 strain

图16 发酵温度对菌株Basidioascus sp. LG-31酶活性的影响Fig.16 Effect of fermentation temperature on the enzyme production of Basidioascus sp. LG-31 strain
3 讨 论
本研究表明,菌株Basidioascussp.LG-31的培养基最佳成分为蔗糖4%、酵母粉0.3%、NaNO30.4%、KH2PO40.02%、KCl 0.10%、MgSO4·7H2O 0.10%。最佳发酵条件:发酵温度20 ℃、初始pH值7.0、三角瓶装液量125 mL/250 mL、接种量2%、转速200 r/min。
本研究为了提高菌株Basidioascussp.LG-31产低温GOD酶活性,对菌株Basidioascussp.LG-31的最佳产酶条件进行单因素优化。实验结果表明,菌株Basidioascussp.LG-31在不同培养条件下产酶能力有很大差异,蔗糖和酵母粉能够大量诱导菌株产酶;菌株在较低温度下的产酶能力明显强于高温下的产酶能力,这可能与菌株来源和低温GOD特性有关;装液量及转速结果表明,较低的装液量和较高的转速更利于菌株产酶,说明菌株发酵产酶对溶氧量需求较高。经过单因素优化后,酶活性达到435.37 U/mL,相对于优化前的酶活性15.28 U/mL,提高了28.49倍,大大提高了菌株Basidioascussp. LG-31的GOD酶活性。2014年石淑钰等[33]从海洋中筛选获得一株产低温GOD的米氏假单胞杆菌(Pseudomonasmigulae),该菌株所产GOD最适催化温度为20 ℃,10 ℃时仍具有较高酶活性,最适pH为7.0,但酶活性最高只有25 U/mL;高齐霖等[34]从海泥中筛选出6株产GOD菌株,其中一株霉菌酶活性最高,为0.904 U/mL;胡善松等[35]从海洋中筛选出一株产GOD柠檬酸杆菌属(Citrobacterssp.)菌株,并对其进行诱变选育和克隆表达,在大肠埃希菌中克隆表达后,重组GOD最适作用温度为25 ℃,酶活性2.04 U/mL;王一茜等[36]以海泥为样品,筛选得到一株产低温GOD的酵母菌WYQ 23,该GOD最适作用温度为20 ℃,并对该菌株产酶进行了单因素发酵条件优化,优化后酶活性达1.67 U/mL。与以往报道的产GOD的菌株相比,菌株Basidioascussp.LG-31在温度和酶活性方面有明显优势,尤其是经过单因素优化后,酶活性明显提高。接下来本课题组(辽宁省海洋微生物工程技术中心)会在本研究的基础上,对菌株Basidioascussp.LG-31产GOD酶活性进行响应面优化,系统性地完成产酶条件优化,以进一步提高产GOD酶活性,并对其发酵罐扩大培养,逐渐进行工业化生产。
GOD在氧气存在的条件下可催化葡萄糖转化,并生成H2O2,从而将巯基(-SH)氧化生成二硫键(-S-S-),增强面粉强度,使其口感更筋道[37-38]。GOD作为一种新型的饲料添加剂,进入动物体内,可以起到帮助分解葡萄糖的作用。催化过程中消耗氧气,降低肠道内氧气含量,抑制好氧病原微生物生存及繁殖,从而维持肠道菌群平衡,有效地提高动物免疫力;生成的葡萄糖酸和H2O2,能够调节畜禽肠道pH,有助于食物的消化吸收[39-40]。本研究中的菌株Basidioascussp. LG-31分离自海洋,因此具有耐低温特性,也可有效解决上述GOD研究现存问题。菌株Basidioascussp. LG-31的成功分离不仅对GOD资源库尤其是酵母来源的GOD的扩充具有重要意义,也为低温GOD的工业化生产及应用提供重要的参考。此外,菌株Basidioascussp.LG-31所产的GOD在低温下仍有很高酶活性,可降低工业生产成本,具有开发前景和应用潜力。
猜你喜欢
阅读(快乐英语中年级)(2022年5期)2022-06-03
中国食用菌(2021年10期)2021-11-04
江西水产科技(2019年3期)2019-09-10
天津农业科学(2019年4期)2019-05-31
中国酿造(2019年4期)2019-05-09
食品工业科技(2019年24期)2019-02-18
农产品加工(2018年21期)2018-11-30
山东工业技术(2016年15期)2016-12-01
中国酿造(2016年12期)2016-03-01
应用海洋学学报(2015年3期)2015-11-22
