
贵州部分洞穴裸灶螽的鉴定及其遗传多样性分析
2022-03-24 08:58吕小艳李斌强何诗远何小涛王野影
环境昆虫学报 2022年1期
吕小艳,李斌强,何诗远,何小涛,王野影*
(1.贵州师范大学生命科学学院,贵阳 550025; 2. 贵州师范大学国家林业局西南喀斯特山地生物多样性保护重点实验室,贵阳 550025)
贵州省拥有丰富的喀斯特生态系统,故而形成众多的喀斯特洞穴地貌类型。洞穴有别于地表,是一个相对封闭的生态环境(王剑等,2000),因此,有着数量庞大的节肢类动物生活在洞穴中。近年来,有大量关于贵州不同喀斯特地区洞穴动物的群落结构的研究(杨卫诚等,2013;梁盛等,2015;张广等,2015;温清,2017),也有针对洞穴贝类、鱼类、以及蜘蛛类动物进行的新种描述(黎道洪等,2003b;陈会明等,2008;郭轩等,2016;杨自忠等,2018;周江,2019),还有针对螺类的多样性研究(黎道洪等,2003a)。昆虫在多数大陆和淡水生态系统中有着很高的多样性和丰富度,其对环境变化具有高度敏感性,故而成为保护和检测研究的模式生物以及生物多样性研究中的指示器,如蚂蚁被用作研究栖息地干扰程度的生物指示物种(Wallaceetal., 1998;王野影,2015)。驼螽科Rhaphidophoridae在世界范围内广泛分布,包含了大量的洞穴适应物种和少量的地表栖息物种(Allegruccietal., 2005)。洞穴灶螽是很多洞穴内的优势类群,是研究自然选择、进化发展生物学、适应性进化和生态环境评估的理想材料。
裸灶螽亚属Tachycines(Gymnaeta)隶属于直翅目Orthoptera驼螽科Rhaphidophoridae疾灶螽属Tachycines,俄罗斯学者Adelung在研究古北界的沙螽时,建立了疾灶螽属Tachycines和裸灶螽亚属Gymnaeta,1992年以前,很多学者一直采用将裸灶螽亚属(Adelung, 1902)归属于疾灶螽属的分类系统(Karny, 1934),1992年以后,Gorochov将裸灶螽亚属从疾灶螽属转至芒灶螽属Diestrammena,并将疾灶螽属认定为芒灶螽属的一个亚属(Gorochov, 1992; 1998)。在2018年的修订中,疾灶螽属被重新提升到属水平,并将裸灶螽亚属重新归属于疾灶螽属,同时,将两个亚种Tachycines(Gymnaeta)ferecaecaGorochov, Rampini & Di Russo, 2006和Tachycines(Gymnaeta)ferecaecaproximaGorochov, Rampini & Di Russo, 2006提升为种水平,将栗色裸灶螽Tachycines(Gymnaeta)aspesRampini & Di Russo, 2008异名为近裸灶螽Tachycines(Gymnaeta)proxima(Qinetal., 2018)。到2009年,全世界报道裸灶螽亚属物种共18种,有16种(包括亚种)来自中国(Zhang & Liu, 2009),其中在贵州分布有5个种和1个亚种(Gorochovetal., 2006;Rampinietal., 2008)。截止到目前,共报道疾灶螽属物种共66种,其中裸灶螽亚属共包含46种(Zhuetal., 2020),贵州分布裸灶螽亚属物种19种。2018年以来,贵州陆续报道疾灶螽属新种高达12种(Qinetal., 2018; Qinetal., 2019; Huangetal., 2019; Fengetal., 2019; Fengetal., 2020; Zhuetal., 2020),充分说明了贵州喀斯特洞穴疾灶螽属物种多样性丰富。随着分子系统学被广泛用于探究系统发育关系、种群历史等各个方面的研究(Yang & Rannala, 2012),部分研究者针对驼螽科物种的系统发育关系开展了分子系统学研究(Valerioetal., 2013; 秦艳艳,2020),但都是从科水平开展的研究,而对于贵州广泛分布的裸灶螽亚属而言,尚未开展遗传多样性研究,缺乏分子方面的数据。
近些年来,随着旅游业的发展,喀斯特洞穴逐渐成为景观旅游开发的对象,原本处于相对稳定和相对封闭状态的洞穴,在被开发后增加了灯光照明和人为活动干扰,从而对洞穴动物造成一定的影响(邓亚东等,2011)。本研究拟通过传统分类方法和分子分类方法相结合,对贵州省部分地区喀斯特洞穴裸灶螽进行物种鉴定,探讨该物种在贵州喀斯特洞穴的分布情况,对洞穴生物进行准确鉴定,有利于增强人们对洞穴生物的认识,在部分濒危物种未灭绝前给予保护。通过构建裸灶螽物种的系统进化树,探讨种间亲缘关系,为后续物种形态鉴定和分类提供参考依据;通过对裸灶螽物种的遗传多样性分析,揭示人为活动干扰和洞穴开发情况对这类生物指示种的影响,从而为洞穴的保护和合理开发提供建议。因此,本研究在喀斯特区洞穴生物物种分类鉴定、分子进化、分子系统学及谱系地理学等方面均具有重要的实践价值和理论意义。
1 材料与方法
1.1 研究对象
2017-2019年间,从贵州省4个地区的8个喀斯特洞穴(黔西南州万峰林邱家洞、熊洞和白龙洞,毕节市大方县关牛洞、摩天大洞和黔西县水牛洞,黔南州长顺县磨谢村菜地洞,铜仁市德江县龙桥村大洞)洞内随机采集裸灶螽共97号标本,将其浸泡于无水乙醇中保存于贵州师范大学生命科学学院重点实验室,详细信息见表1。
1.2 标本形态鉴定
对裸灶螽进行形态鉴定,具体方法参照秦艳艳等的研究(Qinetal., 2019;秦艳艳,2020; Zhuetal., 2020)。以生殖器为主要鉴定特征,借助复眼是否有退化,单眼是否缺失或分离,后足胫节背面刺数作为辅助鉴定特征。首先,将浸泡在无水乙醇中的标本进行回软。然后,在解剖镜下,将回软虫体的腹部末端用解剖针打开,将生殖器与腹部的膜质结构切断,轻轻拉出生殖器,利用解剖镜观察雄性生殖器,雌性标本直接在解剖镜下观察下生殖板。最后,对生殖器进行拍照和特征描绘。
将拍得的标本生殖器照片与裸灶螽亚属的模式种Gymnaetaberezowskii(Adelung, 1902a; Adelung, 1902b)进行生殖器比对,其中主要参照卓氏裸灶螽Tachycines(Gymnaeta)zorzini(Rampinietal., 2008; 张丰,2010; Qinetal., 2019; Zhuetal., 2020)、拉林裸灶螽Tachycines(Gymnaeta)solidus(Fengetal., 2019; 秦艳艳,2020)、实心裸灶螽Tachycines(Gymnaeta)solidus(Gorochovetal., 2006; 温清,2018; Qinetal., 2019; 秦艳艳,2020)、洞穴裸灶螽Tachycines(Gymnaeta)cavernus(Jaioetal., 2008; Zhang & Liu, 2009; Qinetal., 2019; 秦艳艳,2020)、波氏裸灶螽Tachycines(Gymnaeta)borutzkyi(Gorochov & Kireichuk, 1994; 张丰,2010; Qinetal., 2019; 秦艳艳,2020)、伪裸灶螽Tachycines(Gymnaeta)fallax(Zhang & Liu, 2009; Qinetal., 2019; 秦艳艳,2020)、陈氏裸灶螽Tachycines(Gymnaeta)chenhui(Rampinietal., 2008; Qinetal., 2019; 秦艳艳,2020)和拉脱裸灶螽Tachycines(Gymnaeta)latellai(Rampinietal., 2008; Qinetal., 2019; 秦艳艳,2020)的生殖器特征及形态特征进行分类和鉴别。
1.3 标本分子鉴定
1.3.1DNA提取
取裸灶螽标本的后足股节,采用盐析法进行DNA的提取(马婷婷,2011),使用分光光度计检测DNA提取纯度(OD值:1.8~2.0)和浓度,DNA溶液于-20℃冰箱保存待用。
1.3.2PCR扩增及测序
利用由生工生物工程(上海)股份有限公司合成的COI基因序列通用引物Lco1490和Hco2198 (Folmeetal., 1994)进行PCR扩增,普通PCR反应体系为:Taq PCR Master Mix (2X,blue dye)12.5 μL,上下游引物各1 μL (10 μM),DNA模板1 μL (10~100 ng/μL),加ddH2O至25 μL。采用95℃预变性3 min,95℃变性30 s,40℃复性30 s,72℃延伸1 min,重复36个循环,72℃再延伸8 min程序进行扩增。利用1%琼脂糖凝胶电泳检测PCR结果,把具有目标条带的PCR扩增产物直接送至生工生物工程(上海)股份有限公司进行单向测序。
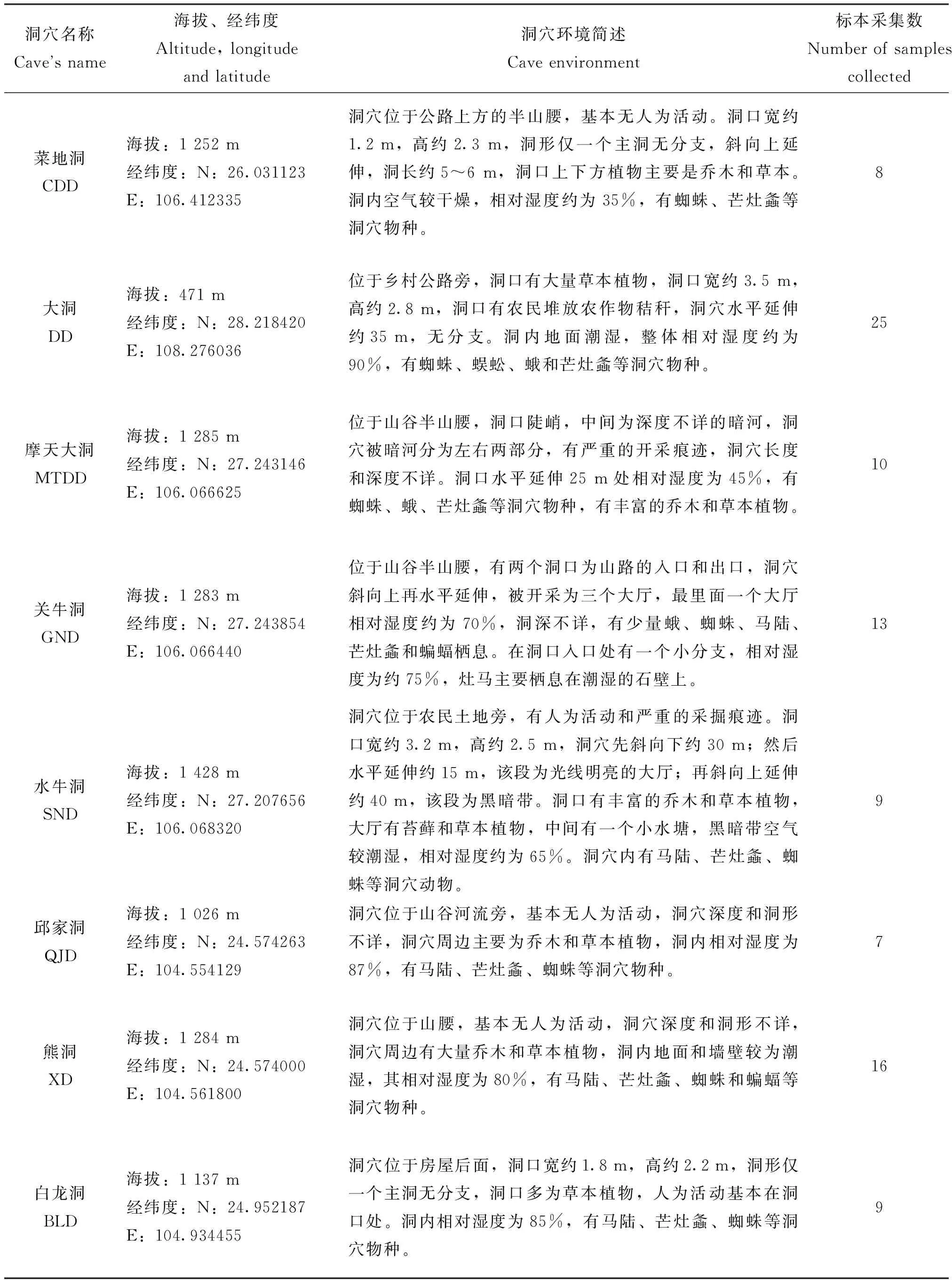
表1 样品采集信息及洞穴描述
1.3.3测序数据处理与分析
首先将测序所得峰图文件使用BioEdit v7.0.5软件(Hall, 1999)进行检验,其序列用NCBI数据库(https://blast.ncbi.nlm.nih.gov/Blast.cgi)进行Blast同源相似性比对,初步判断该序列的来源物种。将均为单峰的序列结合从GenBank数据库中下载的Diestrammenasp.和疾灶螽属的COI序列(表2),全部整理到同一个txt文件中,利用BioEdit v7.0.5将txt文件中所有序列两端比对对齐并保存为Fasta文件。然后利用MEGA 7.0软件(Kumaretal., 2016)以驼螽科的Ceuthophilusgracilipes、Hadenoecuscumberlandicus和Euhadenoecusinsolitus3个物种(焦忠久,2010)作为外群,采用邻接法(NJ),选择Bootstrap method,其他默认设置,1 000次重复构建系统发育树。贝叶斯系统发育分析采用MrBayes 3.2.1软件(Fredriketal., 2012),利用MCMC方法,选择GTR模型,以Ceuthophilusgracilipes为外群共运行300万代,直至两个独立分析的相似度小于0.01,结束运算,使用Figtree v1.4.3软件(Rambaut & Drummond, 2012)进行进化树的编辑和查看。Adobe Illustrator CS4进行进化树的美化。
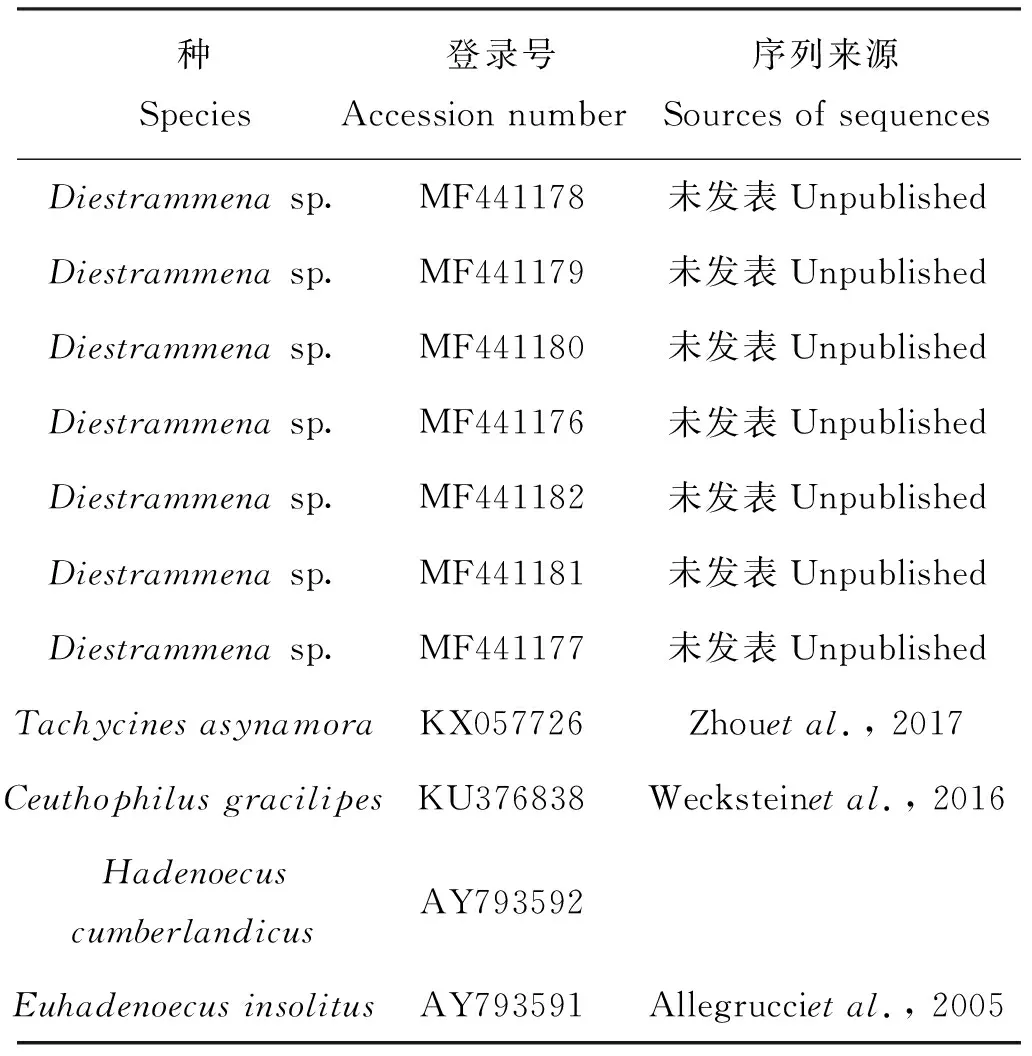
表2 芒灶螽属、庭疾灶螽及外群COI序列GenBank检索号
利用MEGA 7.0软件采用p-距离法,bootstrap评估变异,重复1 000次,对97号裸灶螽标本种间遗传距离进行分析;使用DnaSPv6软件(Rozasetal., 2017)对已鉴定的物种进行COI基因的变异位点(Variable sites),简约信息位点(Parsimony informative sites)、单倍型H及单倍型多样性Hd和核酸多样性Pi的分析。
2 结果与分析
2.1 裸灶螽形态鉴定
通过对97号裸灶螽标本生殖器及形态特征进行鉴定,获得卓氏裸灶螽(图1-a,图1-b)、拉林裸灶螽(图1-c,图1-d)、洞穴裸灶螽(图1-e)、实心裸灶螽(图1-f)、波氏裸灶螽(图1-g,图1-h)、伪裸灶螽(图1-i,图1-j)、陈氏裸灶螽(图1-k,图1-l)和拉脱裸灶螽(图1-m,图1-n)共8个种,均属于驼螽科疾灶螽属裸灶螽亚属,由于有的裸灶螽标本为若虫,生殖器未发育成型,或同种标本数量较少只有一个雌性标本,无法通过形态鉴定到具体物种,在本研究中统称为待定种(表3)。由表3可知,来自不同地区的洞穴裸灶螽物种,其种类各不相同,黔南地区8号标本鉴定为拉脱裸灶螽和2个待定种;铜仁地区25号标本鉴定为拉林裸灶螽、洞穴裸灶螽、实心裸灶螽和波氏裸灶螽4个种,以及1个待定种;毕节地区32号标本鉴定结果全为卓氏裸灶螽;黔西南地区32号标本鉴定为伪裸灶螽、陈氏裸灶螽和一个待定种。由邱家洞(QJD)7号标本共鉴定出3个种,而熊洞(XD)16号标本共鉴定出2个种,说明裸灶螽标本数与物种数不存在相关性。
2.2 分子系统发育分析
DNA提取纯度在260/280波长下,OD值均处于1.8~2.0之间,PCR产物条带大小在650 bp左右,其测序结果序列在NCBI中与庭疾灶螽Tachycinesasynamora的同源性相似比均大于87%,表明测序结果可信,序列可用于后续分析。
在分子系统发育进化树上(图2),97条COI序列分为Clade A、Clade B、Clade C、Clade D、Clade E和一个待定种Tachycines(Gymnaeta) sp4. 6个支系,除Clade B分支上NJ支持率为90%外,其余分支NJ支持率均大于95%,BI后验率为均大于0.98,说明各分支具有很高的可信度。根据形态鉴定结果的标本序号对应,Clade A包括卓氏裸灶螽和拉林裸灶螽2个种;Clade B包括3个待定种Tachycines(Gymnaeta) sp1.、Tachycines(Gymnaeta) sp2.和Tachycines(Gymnaeta) sp3.以及洞穴裸灶螽、实心裸灶螽和波氏裸灶螽3个种;Clade C包括伪裸灶螽1个种;Clade D包括陈氏裸灶螽1个种;Clade E包括拉脱裸灶螽1个种。
从进化树上可知,卓氏裸灶螽和拉林裸灶螽聚为一支,然后再与洞穴裸灶螽、实心裸灶螽和波氏裸灶螽聚在一起,且BI后验率为1。Tachycines(Gymnaeta) sp3.与洞穴裸灶螽聚在一起,且BI后验率为0.98,说明Tachycines(Gymnaeta) sp3.也为洞穴裸灶螽。Clade B支系上有很多小分支,已知标本的体色均为体黄(深)褐色,杂有暗(黄)褐色条纹,且雌性下生殖板均近三角形,雄性外生殖器背骨片均近“H”形,因此,推测该支系可能存在复合种。同时,从GenBank中下载的已知Diestrammenasp.序列,在进化树上位于已知庭疾灶螽Tachycinesasynamora序列的内测,推测所下载的GenBank中Diestrammenasp.序列可能属于疾灶螽属Tachycines。
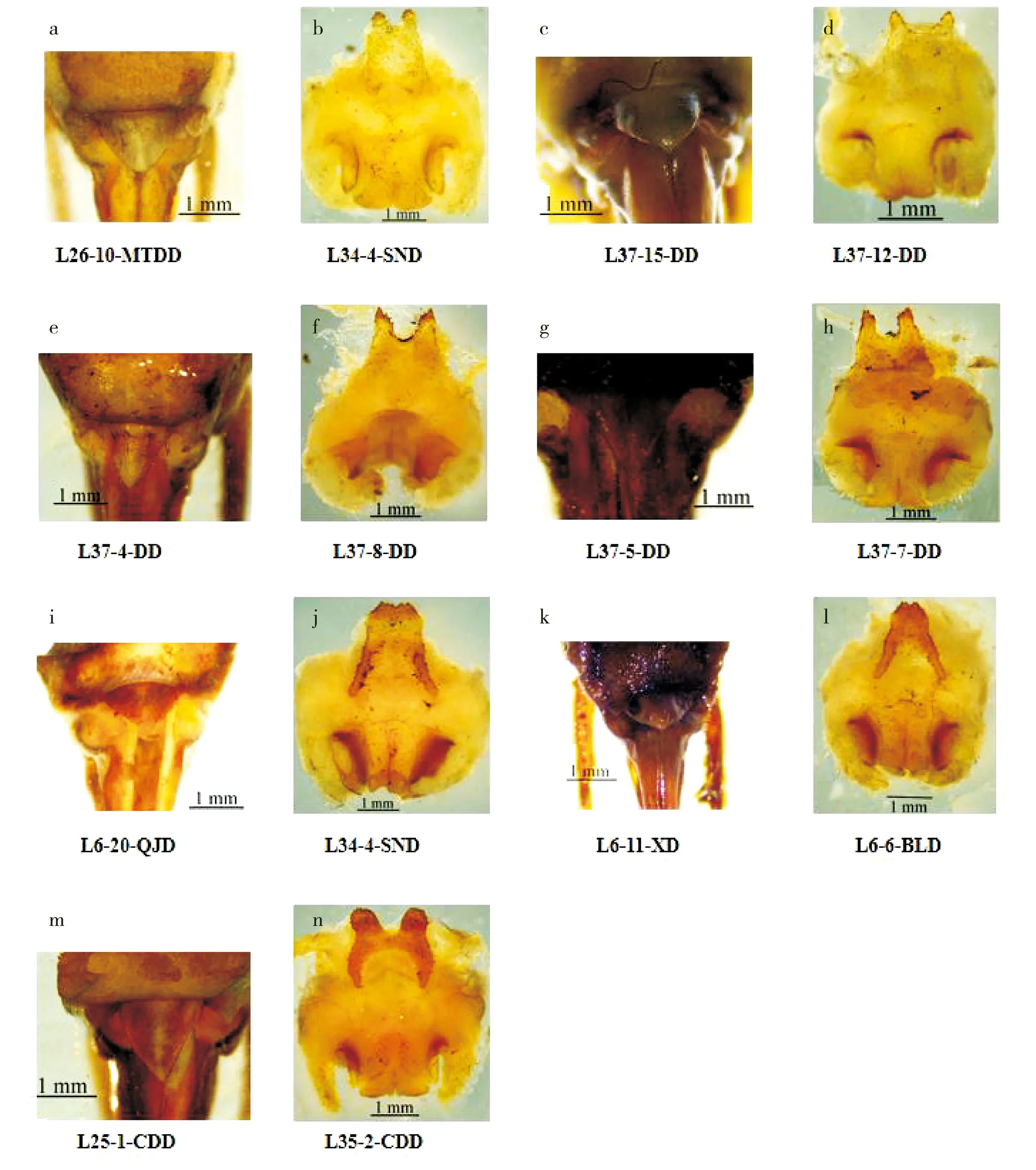
图1 裸灶螽亚属形态鉴定图Fig.1 Morphological identification of Tachycines (Gymnaeta)注:卓氏裸灶螽:a,雌性下生殖板腹面观;b,雄性生殖器背面观;拉林裸灶螽:c,雌性下生殖板腹面观;d,雄性生殖器背面观;实心裸灶螽:e,雌性下生殖板腹面观;洞穴裸灶螽:f,雄性生殖器背面观;波氏裸灶螽:g,雌性下生殖板腹面观;h,雄性生殖器背面观;伪裸灶螽:i,雌性下生殖板腹面观;j,雄性生殖器背面观;陈氏裸灶螽:k,雌性下生殖板腹面观;l,雄性生殖器背面观;拉脱裸灶螽:m,雌性下生殖板腹面观;n,雄性生殖器背面观。Note: Tachycines (Gymnaeta) zorzini: a, Female subgenital plate, ventral view; b, Male genitalia, dorsal view; Tachycines (Gymnaeta) lalinus: c, Female subgenital plate, ventral view; d, Male genitalia, dorsal view; Tachycines (Gymnaeta)solidus: e, Female subgenital plate, ventral view; Tachycines (Gymnaeta) cavernus: f, Male genitalia, dorsal view; Tachycines (Gymnaeta) borutzkyi: g, Female subgenital plate, ventral view; h, Male genitalia, dorsal view; Tachycines (Gymnaeta) fallax: i, Female subgenital plate, ventral view; j, Male genitalia, dorsal view; Tachycines (Gymnaeta)chenhui: k, Female subgenital plate, ventral view; l, Male genitalia, dorsal view; Tachycines (Gymnaeta)latellai: m, Female subgenital plate, ventral view; n, Male genitalia, dorsal view.
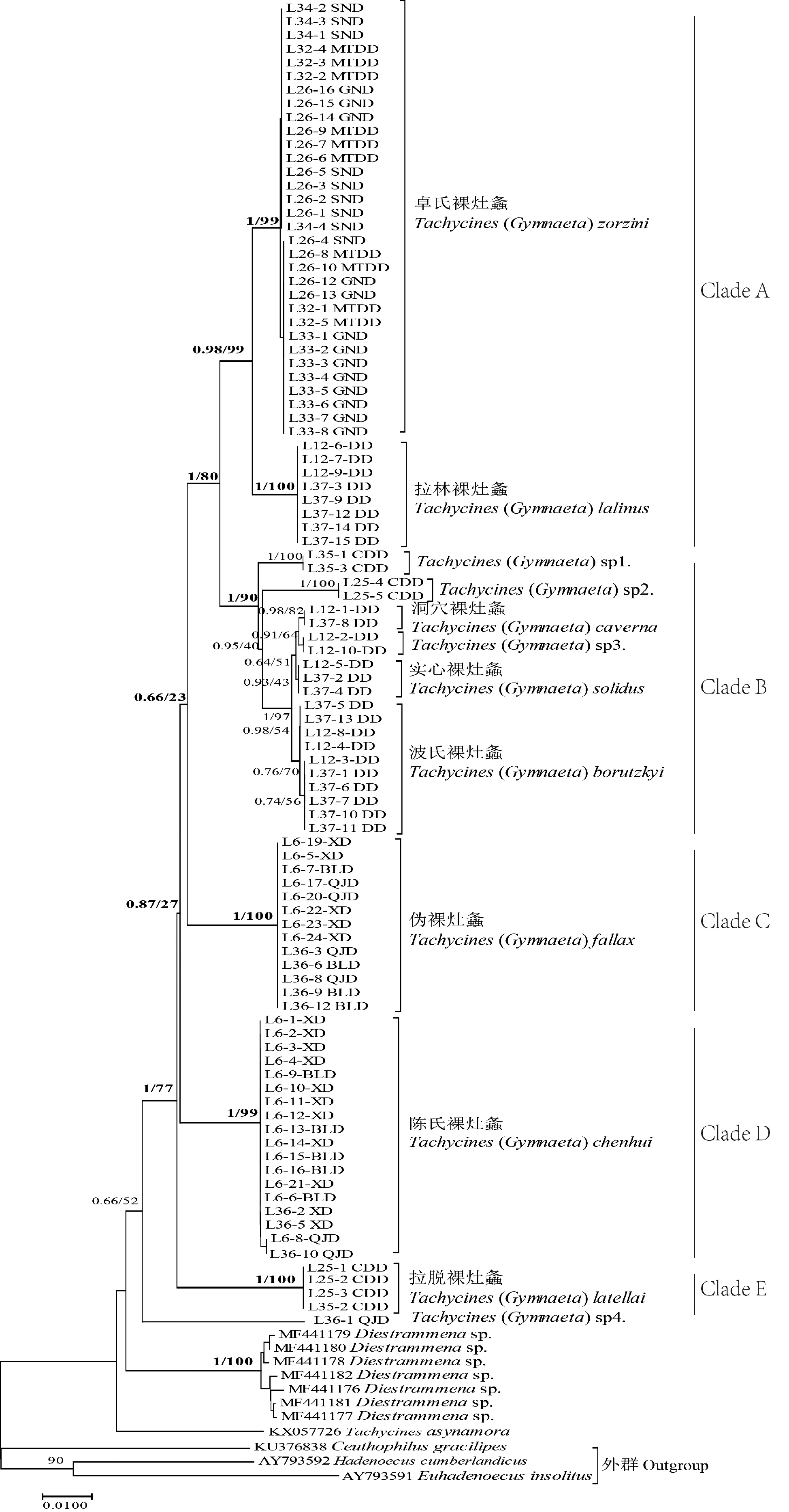
图2 基于106条COI序列的系统发育进化树Fig.2 Phylogenetic tree based on 106 COI sequences 注:树枝上的数字代表分支的后验概率BI/NJ。Note:The number on the branch represents the posterior probability of the branch BI/NJ.
2.3 种间遗传距离分析
MEGA 7.0软件遗传距离数据输出为左下三角和右上三角两个矩阵,左下三角表示遗传距离,右上三角表示遗传分化系数。在2.2分析中得出Tachycines(Gymnaeta)sp3.同为洞穴裸灶螽,因此,将二者合为洞穴裸灶螽进行分析。由表4可知,裸灶螽亚属的种间遗传距离在0.0055至0.1366之间变化,其中最大遗传距离0.1366为Tachycines(Gymnaeta) sp2.与Tachycines(Gymnaeta)sp4.之间,最小遗传距离0.0055为洞穴裸灶螽与实心裸灶螽之间。除此,洞穴裸灶螽与波氏裸灶螽之间的距离,以及波氏裸灶螽与实心裸灶螽之间的遗传距离也很小,均为0.0078。因此,洞穴裸灶螽、波氏裸灶螽和实心裸灶螽之间的种间遗传距离最小,亲缘关系最近。
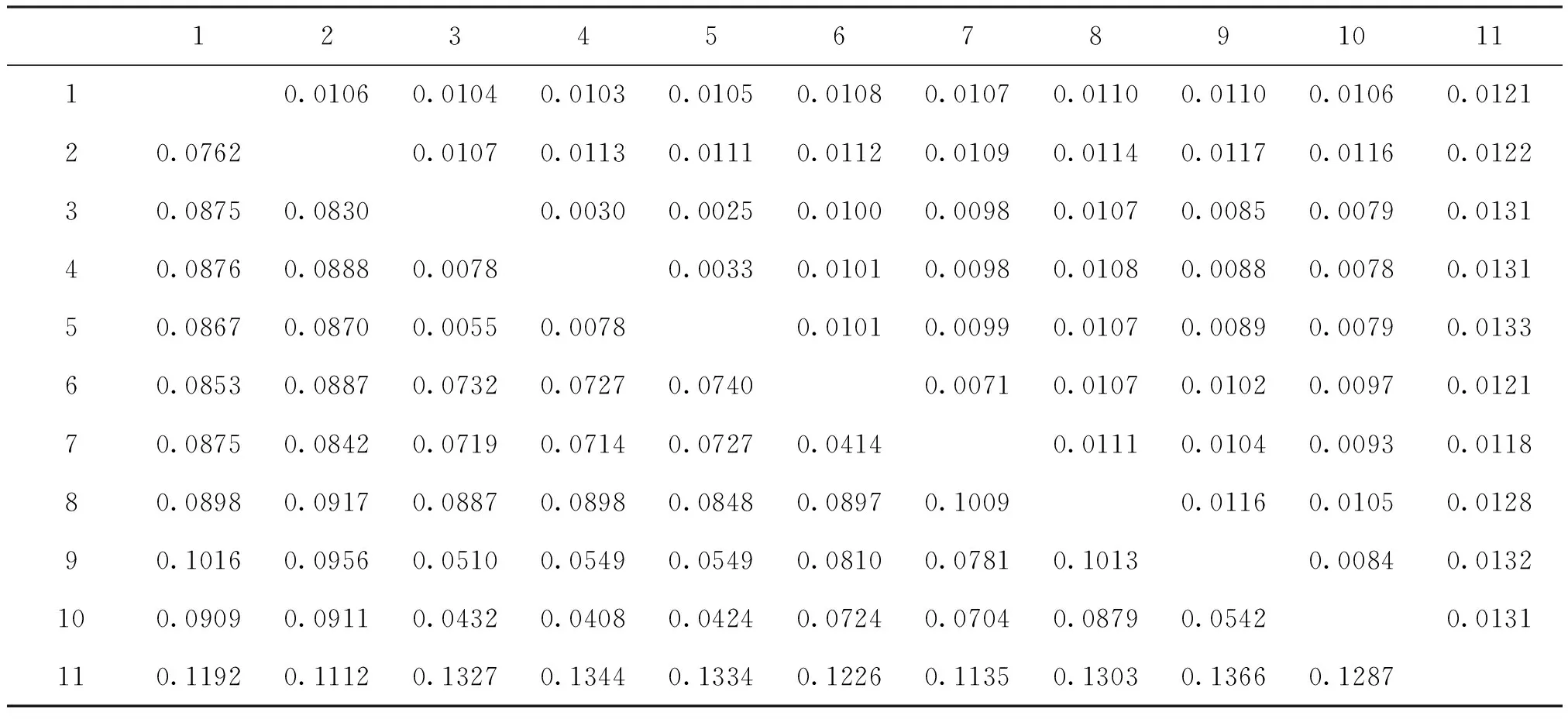
表4 裸灶螽亚属的种间遗传距离
2.4 遗传多样性分析
从物种遗传多样性指数统计表(表5)中可看出,同一洞穴中物种遗传多样性为:大洞中波氏裸灶螽和洞穴裸灶螽的遗传多样性最高;菜地洞中Tachycines(Gymnaeta) sp2.的遗传多样性最高;白龙洞和熊洞中伪裸灶螽的遗传多样性最高;邱家洞中陈氏裸灶螽的遗传多样性最高。不同洞穴同一物种遗传多样性为:伪裸灶螽和陈氏裸灶螽在邱家洞中遗传多样性最高,在熊洞中遗传多样性最低;卓氏裸灶螽在水牛洞中遗传多样性最低。
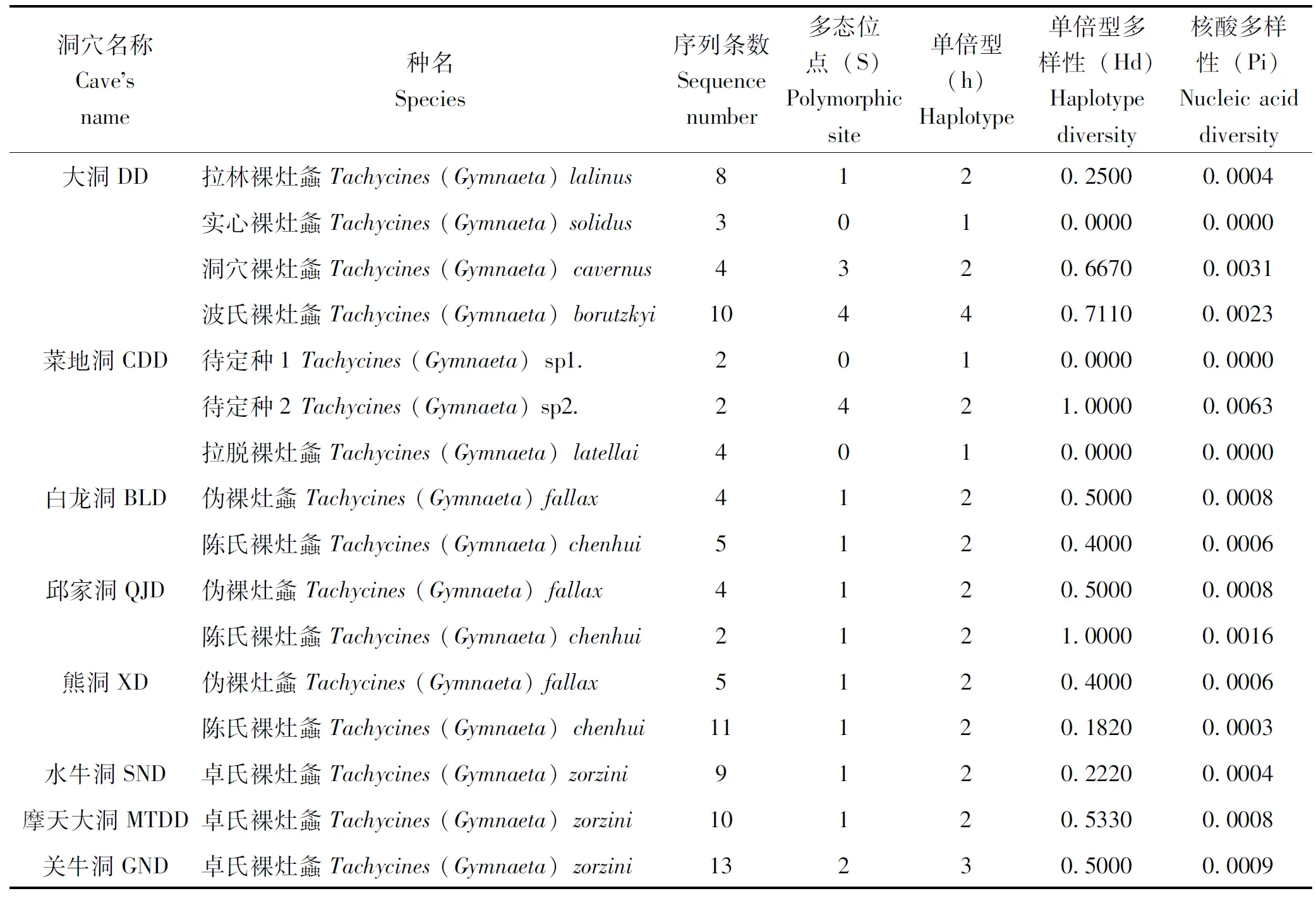
表5 各洞穴中物种遗传多样性指数统计表
所有物种遗传多样性由高至低分别为:陈氏裸灶螽、波氏裸灶螽、洞穴裸灶螽、卓氏裸灶螽、伪裸灶螽、拉林裸灶螽、实心裸灶螽和拉脱裸灶螽。其中,实心裸灶螽和拉脱裸灶螽的单倍型均为1,其变异位点、单倍型多样性和核酸多样性均为零,除标本数量较少外,还可能存在所采集的标本均来自相同的亲本的情况。
3 结论与讨论
3.1物种鉴定
我国作为驼螽科昆虫的重要产地,吸引不少研究者对其进行分类研究,但大都致力于属的修订和新种的描述,由于不同的分类学者持不同的分类观点,导致分类系统难以统一;有的昆虫在若虫阶段时,难以利用生殖器区分具体的物种,或只通过形态特征进行鉴定容易出现错误,而基于分子条形码的物种鉴定技术则可以弥补其中的不足之处,辅助物种鉴定(林森杰等,2014; 李海涛等,2019),甚至弥补不同发育阶段标本形态鉴定难的问题(Barberetal., 2006; Puillandreetal., 2009),同时,线粒体基因组序列被认为是解决种下阶元到目级阶元最广泛最有效的分子遗传标记(Cameron, 2014),如利用线粒体基因组构建直翅目高级阶元(科、总科)之间的系统发育树探讨其亲缘关系(Songetal., 2015; Zhouetal., 2017)。本研究利用COI基因作为分子标记,成功对裸灶螽亚属物种进行鉴定,说明线粒体DNAbarcoding基因在裸灶螽亚属物种鉴定中具有普适性。在构建的分子进化树上,卓氏裸灶螽与拉林裸灶螽的亲缘关系最近,这与二者在形态鉴定描述上极为相似是一致的(Fengetal., 2019),同时,洞穴裸灶螽、实心裸灶螽和波氏裸灶螽在进化树上聚为一支,且遗传距离最近,三者在形态鉴定描述上也极为相似(秦艳艳,2020),除此,陈氏裸灶螽与拉脱裸灶螽在进化树上的位置,与二者在意大利学者联合COI和ITS1基因对灶螽亚科构建的系统发育树上的位置极其相似(Valerioetal., 2013),说明本研究通过形态结合分子的方法鉴定出的结果具有很高的可信度。鉴定中遗留的4个待定种因其为若虫和缺少雄性标本,而且分子数据库缺乏相关的数据比对,故而也无法通过系统发育鉴定到具体物种,在接下来的研究中,需对菜地洞、大洞和邱家洞进行重复采样,尽可能选择采集成虫,对其进行形态鉴定以确定具体物种。
3.2 裸灶螽分布受地理隔离的影响
伪裸灶螽的模式产地在云南腾冲大蒿坪(Zhang & Liu, 2009),本研究中采自贵州黔西南地区的标本也鉴定出该物种,黔西南地区与云南邻界,可能两地存在基因交流。洞穴裸灶螽的模式产地在湖北通山隐水洞(Jaioetal., 2008; Qinetal., 2019),在湖南也有分布,与模式产地为重庆的波氏裸灶螽(Gorochov, 1998)和模式产地为湖南龙山的实心裸灶螽(Gorochovetal., 2006)很相似,后两种在贵州均有分布(温清,2018;Qinetal., 2019; 秦艳艳,2020),本研究采自贵州铜仁地区的标本也鉴定出洞穴裸灶螽物种,铜仁地区与湖南及重庆邻界,因此,说明地区之间可能存在基因交流。鉴定结果中,来自毕节地区的摩天大洞、关牛洞和水牛洞之间,以及来自黔西南地区的邱家洞、熊洞和白龙洞之间,不同洞穴中存在相同物种,而对于来自毕节、黔西南、黔南和铜仁不同地区洞穴之间不存在相同物种,可能是受贵州“四山八水”造成的地理隔离的影响,也说明裸灶螽活动能力较弱,适合进行洞穴生物起源和进化历史的假说验证研究。
3.3 洞穴环境与裸灶螽遗传多样性的关系
核苷酸多样性Pi用于衡量物种或种群的多态性程度,其数值越大说明物种或种群的遗传多样性就会越高(Avise & Zink, 1988)。从物种遗传多样性结果结合洞穴描述可知,菜地洞位于半山腰,虽洞穴空间较小,但基本无人为活动,没有开采痕迹,因此,遗传多样性最高;邱家洞、大洞、白龙洞和熊洞人为活动干扰少,没有开采痕迹,洞内相对湿度较高,其遗传多样性也较高;关牛洞、水牛洞和摩天大洞虽洞穴空间大,但人为活动干扰频繁,开发痕迹严重,因此,仅发现一种裸灶螽物种,且32条COI基因仅有一个变异位点,说明洞穴的开发对洞内物种遗传多样性以及物种种类的下降有一定的影响。洞穴旅游开发对生态环境影响分析发现,洞穴开发前后洞中蝙蝠、芒灶螽和蜘蛛数量会减少(邓亚东等,2011)。洞栖蝙蝠的多样性研究显示洞穴的旅游开发对蝙蝠的栖息地造成了严重破坏,使得洞栖蝙蝠面临着生存危机(卜艳珍等,2014)。因此,对于洞穴空间较大,物种较丰富的洞穴,应合理开发,尽量减少人为干扰并加强科学管理与保护,避免旅游开发而造成物种灭绝。对于遗传多样性指数为零的物种,如实心裸灶螽和拉脱裸灶螽等,除增加标本数据进一步研究外,也需重视洞穴中该物种的保护。
致谢:非常感谢周旭林师兄在形态鉴定上的指导和帮助。
猜你喜欢
自我保健(2021年10期)2022-01-12
疯狂英语·新阅版(2020年5期)2020-06-12
阅读(快乐英语高年级)(2020年8期)2020-01-08
健康之友·下半月(2019年11期)2019-10-21
信息化建设(2019年7期)2019-09-18
小学生时代·综合版(2017年6期)2017-07-05
消费电子(2017年1期)2017-02-16
办公室业务(2016年11期)2017-01-09
现代养生·下半月(2015年9期)2015-09-28
现代商贸工业(2009年17期)2009-07-06
