
家蚕Fox转录因子在精巢中的表达特征及BmFoxL2在精巢发育中的功能初探
2022-03-24 10:24胡启豪戴玉玲裴梦圆肖妍虹赵丹珲余小强卢玉珍
环境昆虫学报 2022年1期
胡启豪,戴玉玲,裴梦圆,肖妍虹,赵丹珲,余小强,卢玉珍
(华南师范大学生命科学学院,广东省昆虫发育生物学与应用技术重点实验室,广州 510631)
家蚕是我国重要的经济益虫,经过长期的驯化和人工育种,目前已经培育出大批具备优良性状的家蚕品种。遗传杂交是人工育种的关键步骤,其中雄性家蚕精子质量的高低直接影响着后代的性状。精巢作为精子发生的场所,其发育机制的研究对于家蚕遗传育种有着重要的意义。
Fox家族蛋白存在于植物以外的生物类群当中,具有一个约含100个氨基酸的保守结构域,根据不同Fox蛋白的保守性,可分为19个亚族:FoxA~FoxS(Weigeletal., 1989; Kerschneretal., 2014)。不同Fox蛋白已被证明在哺乳动物和昆虫的组织发育和先天免疫等多种生物学过程中发挥关键作用。在黑腹果蝇Drosophilamelanogaster中,FoxN3(CHES-1-like)的过表达会导致精巢形态异常,并抑制精子发生的过程(Yuetal., 2016)。甜菜夜蛾SpodopteraexiguaSeFox蛋白结构与家蚕BmFoxA相似,并且在精巢中有一定水平的表达(Zhaoetal., 2014)。家蚕的BmFoxG-1可能通过调节BmCyclinA等细胞周期基因的表达,从而调节家蚕精巢的发育(胡启豪等, 2021)。Song等通过基因芯片(Microarray)的方法分析发现不同Fox基因在家蚕精巢中都有不同程度的表达(Songetal., 2015)。这些研究都说明Fox基因的功能可能与精巢发育相关,但其具体作用机制仍不清楚。
本研究通过qRT-PCR检测了不同BmFox基因在家蚕5龄第5天(L5D5)幼虫和预蛹期精巢中的表达水平,结果发现BmFoxL2亚族基因在两个时间点均有高水平表达。本研究利用生物信息学方法对家蚕FoxL2亚族蛋白的结构进行了分析,并进一步分析BmFoxL2-1和BmFoxL2-2在家蚕不同发育阶段精巢中的表达情况以及在Bm12中的表达定位。最后,在家蚕Bm12细胞中分别过表达BmFoxL2-1和BmFoxL2-2蛋白,并分析了BmVasa等生殖和细胞周期相关基因的表达变化。本研究的结果初步说明BmFoxL2亚族蛋白参与了家蚕精巢发育及精子发生的过程。
1 材料与方法
1.1 材料和主要试剂
本研究使用的大造家蚕品种由广东省蚕业技术推广中心提供,家蚕的培养条件参考的方法胡启豪等( 2021)。家蚕Bm12细胞、大肠杆菌DH5α和细胞过表达载体IE1-pEGFP-N1等均为本实验室保存材料。
Trizol总RNA提取试剂购自中国艾科瑞生物公司;反转录试剂盒和qRT-PCR试剂购自康为世纪生物技术有限公司;细胞转染试剂购自Qiagen公司;引物由北京擎科新业生物技术有限公司合成;去内毒素质粒小量抽提试剂盒购自广州美基生物科技有限公司;实验所用其它试剂均为国产分析纯试剂。
1.2 生物信息学分析
从家蚕基因组数据库SilkDB中获得BmFoxL2-1(BGIBMGA000635)和BmFoxL2-2(BGIBMGA000838)基因及蛋白质序列。蛋白质理化性质预测通过ExPASy进行,磷酸化位点及糖基化位点利用Cbs网站进行分析,蛋白结构域利用SMART进行分析。
1.3 不同发育阶段家蚕精巢cDNA制备
收集5龄第3天(L5D3)幼虫至成虫第3天的雄性家蚕,在PBS缓冲液中进行解剖并取出精巢。采用Trizol法提取总RNA。逆转录反应按照康为世纪HiFiScript gDNA removal RT Master Mix 说明书进行,cDNA保存于-20℃。
1.4 不同Fox基因在家蚕不同发育阶段精巢中的表达量检测
以稀释后的cDNA为模板,采用qRT-PCR方法分析目标基因的表达情况,以家蚕rp49作为内参基因。qRT-PCR循环条件如下:95°C 5 min;95°C 10 s,60°C 30 s,40个循环。本研究所用引物序列见表1。

表1 实验所用引物的序列
1.5 BmFoxL2-1/-2细胞过表达重组质粒的构建
根据BmFox2-1/-2的cDNA序列,设计上下游引物(表1),克隆方法参考的方法胡启豪等(2021)。利用SacⅠ和BamHⅠ对目的片段及pEGFP-N1-GFP载体进行双酶切,DNA凝胶回收后用T4连接酶将两者连接构建重组载体,经PCR与双酶切验证后转化进大肠杆菌DH5α中保存。采用去内毒素质粒抽提试剂盒抽提质粒供转染使用。
1.6 BmFoxL2-1/BmFoxL2-2在 Bm12细胞株的过表达及细胞定位分析
将Bm12细胞用新鲜培养基悬浮,分装至6孔板中,并补足培养基至每孔2 mL,当每孔细胞密度达到80%以上时,参考Qiagen转染试剂说明书的方法将BmFoxL2-1-EGFP和BmFoxL2-2-EGFP质粒转染Bm12细胞。
转染48 h后,去除细胞培养液,加入适量PBS清洗细胞。用4%多聚甲醛固定细胞,用1‰ PBT(含1‰ Triton X-100的PBS)对固定后的细胞进行通透。往细胞中加入适量浓度的DAPI对细胞进行染色,结束后用1‰ PBT清洗多余的DAPI。制片后于激光共聚焦显微镜下分析BmFoxL2-1-EGFP和BmFoxL2-2-EGFP在细胞中的定位。
2 结果与分析
2.1 不同Fox基因在家蚕精巢中的表达
家蚕存在有核与无核两种形态的精子,分别在幼虫阶段和预蛹期形成,因此本研究收集了5龄第5天(L5D5)幼虫和预蛹期(PP)家蚕精巢。定量分析显示,在两个时期的家蚕精巢中,BmFoxL2-2的表达水平均显著高于其它BmFox基因,FoxL2亚族的BmFoxL2-1和FoxO亚族的BmFoxO-1/BmFoxO-2的表达水平次之(图1)。因此,在后续的研究中将重点分析BmFoxL2-1和BmFoxL2-2在精巢发育中的作用。

图1 不同BmFox基因在幼虫和预蛹期家蚕精巢中的表达Fig.1 Expression of different BmFox genes in the testis of Bombyx mori larvae and prepupae注:A,5龄第5天幼虫;B,预蛹。不同字母表示差异性显著(单因素方差分析,Turkey多重比较,P<0.05)。Note: A, The day 5 of 5th instar larvae; B, Prepupae. Different letters indicated significant difference between the two groups (One way ANOVA: Tukey’s HSD tests, P<0.05).
2.2 BmFoxL2蛋白的结构及理化性质分析
为进一步了解BmFoxL2的功能,本研究对BmFoxL2蛋白的理化性质进行分析。结果显示BmFoxL2-1和BmFoxL2-2蛋白的分子量分别为20.35 kDa和24.52 kDa,等电点分别为9.85和5.87,BmFoxL2-1蛋白的磷酸化和糖基化位点都高于BmFoxL2-2(表2)。利用SMART网站对2个BmFoxL2亚族蛋白的结构预测,结果显示均含一个Forkhead结构域(图2-A),并且Forkhead结构域的序列与其它昆虫以及人类FoxL2亚族蛋白的高度保守(图2-B)。

图2 BmFoxL2-1和BmFoxL2-2蛋白结构、序列和定位分析Fig.2 Structural analysis, sequence aglinment and cellular localization of BmFoxL2-1 and BmFoxL2-2注:A,蛋白结构预测;B,氨基酸序列比对;C,细胞定位分析。Note: A, Structural analysis; B, Sequence aglinment; C, Cellular localization.

表2 BmFoxL2-1和BmFoxL2-2蛋白理化性质分析
为了进一步确认BmFoxL2-1/BmFoxL2-2的亚细胞定位,将构建好的BmFoxL2-1-EGFP和BmFoxL2-2-EGFP质粒转染家蚕Bm12细胞株(图2-C)。结果显示BmFoxL2-1定位于细胞核,BmFoxL2-2在细胞核与细胞质中都有分布,这可能是由于BmFoxL2-1和BmFoxL2-2上所含的磷酸化和O-糖基化位点数目不一所导致的。以上结果说明BmFoxL2-1和BmFoxL2-2的功能可能存在差异。
2.3 BmFoxL2亚族基因在精巢中的表达
为进一步分析BmFoxL2亚族基因的功能,本文检测了两个FoxL2亚族基因在5龄第5天家蚕幼虫精巢和卵巢中的表达变化,发现两个BmFoxL2亚族基因在精巢中的表达均显著高于卵巢(图3-A)。由于精巢是精子发生的主要场所,因此本研究对5龄第5天家蚕幼虫精巢的精巢膜和精巢内容物进行分离。对精巢膜和内容物的基因表达检测显示两个BmFoxL2亚族基因在精巢内容物中的表达水平均显著高于精巢膜(图3-B),说明BmFoxL2蛋白可能参与精子发生。
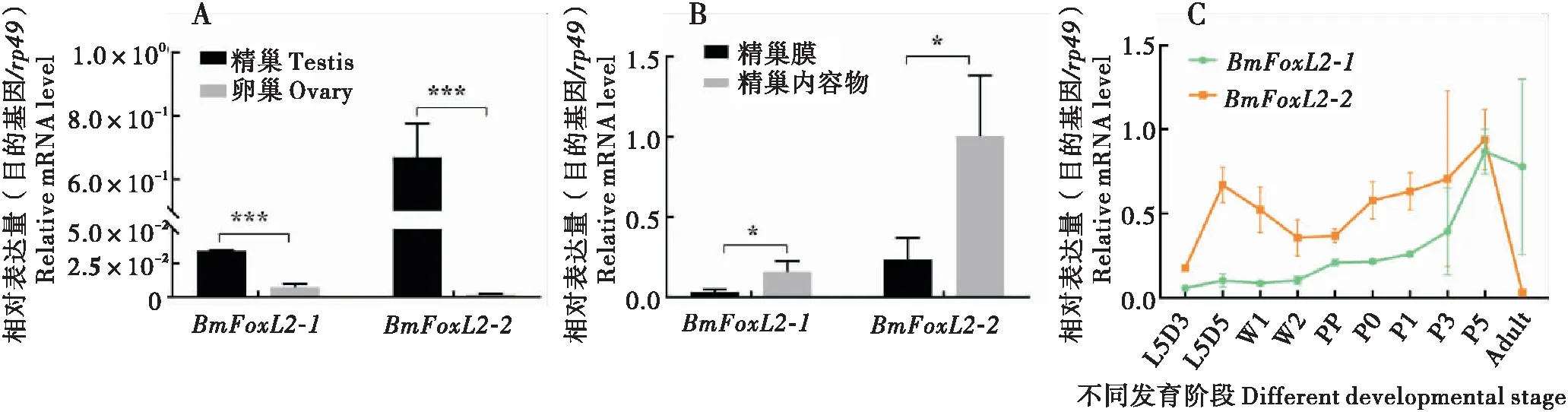
图3 BmFoxL2-1 和BmFoxL2-2基因在家蚕中的表达Fig.3 Expression of BmFoxL2-1 and BmFoxL2-2 transcripts in silkworm注:A,BmFoxL2-1和BmFoxL2-2基因在家蚕精巢和卵巢中的表达;B,BmFoxL2-1和BmFoxL2-2基因在精巢内容物与精巢膜中的表达;C,BmFoxL2-1和BmFoxL2-2基因在不同发育阶段的精巢中的表达变化。柱上标有星号表示两组的表达差异显著(*,P<0.05;***,P<0.001;t检验)。Note: A, Expression of BmFoxL2-1 and BmFoxL2-2 transcripts in the testis and ovary of silkworm at day 5 of 5th instar larvae; B, Expression of BmFoxL2-1 and BmFoxL2-2 mRNAs in different parts of the testis at day 5 of 5th instar larvae; C, Expression of BmFoxL2-1 and BmFoxL2-2 transcripts in the testis of silkworm at different developmental stages. The asterisk above the column indicated significant difference between the two groups (*, P<0.05; ***, P<0.001; t-test).
为了分析BmFoxL2基因在精巢中的表达模式,本研究在5龄第3天幼虫到成虫第1天的家蚕精巢中通过qPCR检测两个BmFoxL2基因的表达量。结果显示,BmFoxL2-1的表达水平随着精巢的发育逐渐升高,但在幼虫阶段增速缓慢,而进入蛹期后,上升速度开始加快。BmFoxL2-2的表达水平在5龄幼虫末期(L5D5)达到一个高峰,在变态发育阶段逐渐下降(L5D5-PP),在蛹期阶段(P0-P5)缓慢回升,到了成虫阶段迅速下降。但BmFoxL2-2在不同发育阶段精巢中的表达均高于BmFoxL2-1(图3-C)。该结果进一步暗示BmFoxL2-2可能在有核精子的形成中发挥作用,而BmFoxL2-1和BmFoxL2都可能与无核精子的形成相关。
2.4 BmFoxL2亚族蛋白对精巢生殖细胞发育的影响
细胞周期蛋白与生殖细胞有丝分裂和减数分裂等过程有着密不可分的关系。报道显示BmFoxG-1可能通过调节细胞周期等基因的表达,参与精子发生过程。本研究在Bm12细胞株中过表达了BmFoxL2-1基因,发现细胞周期基因BmCyclinA的表达显著上调,而生殖干细胞发育关键基因BmNanos的表达也发生了显著上调(图4)。而过表达BmFoxL2-2基因后,BmCyclinA、BmCyclinB和BmCyclinB3等细胞周期基因的表达显著上调,BmNanos和BmVasa等生殖干细胞发育关键基因的表达也发生了显著上调(图5)。以上结果说明,两个BmFoxL2亚族蛋白都可能通过调节生殖干细胞的发育及精细胞分裂等过程,影响家蚕精子的发生,而且BmFoxL2-2的调控功能更加显著。

图4 BmFoxL2-1蛋白对精子发生相关基因表达的影响Fig.4 Effect of BmFoxL2-1 overexpression on the expression of genes related to spermatogenesis注:柱上标有星号表示两组的表达差异显著;NS两组的表达差异不显著(*,P<0.05;**,P<0.01;t检验)。Note: Asterisks above the columns indicated significant difference between the two groups, “NS” indicates non-significant between the two groups (*, P<0.05; **, P < 0.01; t-test).

图5 BmFoxL2-2蛋白对精子发生相关基因表达的影响Fig.5 Effect of BmFoxL2-2 overexpression on the expression of genes related to spermatogenesis注:柱上标有星号表示两组的表达差异显著;NS两组的表达差异不显著(*,P<0.05;t检验)。Note: Asterisks above the columns indicated significant difference between the two groups; “NS” indicated non-significant between the two groups (*, P<0.05; t-test).
3 结论与讨论
FoxL2为Fox家族蛋白的一员,被认为是动物性别分化的关键因子。在小鼠中,FoxL2纯合突变可以导致次级卵泡的缺失和卵母细胞的闭锁(Uhlenhautetal., 2009)。在褐飞虱Nilaparvatalugens和沙漠飞蝗Schistocercagregaria中也发现FoxL2在卵巢中有较高水平的表达,并影响卵巢的发育(De Loofetal., 2010; Yeetal., 2017)。尽管也有研究显示FoxL2在泥蟹Scyllaparamamosain等动物精巢中的表达高于卵巢,家蚕Microarray数据也显示FoxL2-2基因在精巢中的表达高于卵巢,但FoxL2蛋白在精巢中的功能仍不清楚(Songetal., 2015; Wanetal., 2021)。本研究通过qRT-PCR实验发现两个家蚕BmFoxL2基因在精巢中的表达水平显著高于卵巢,并且主要在精巢内容物中表达。
在家蚕精子发生过程中,精原细胞可以分化为有核精子与无核精子。在无核精子的辅助下,有核精子与家蚕卵结合形成受精卵(Sahara and Kawamura, 2002; Pereira and Santos, 2015)。家蚕精子的二型分化具有严格的时期特异性,即有核精子在幼虫阶段开始发生,而无核精子的发生则要到预蛹期(PP)才能开始,这些过程受到不同基因的严格调控(Sahara and Kawamura, 2002)。本研究分析了BmFoxL2-1和BmFoxL2-2在不同发育阶段家蚕精巢中的表达变化,结果发现BmFoxL2-1在幼虫精巢中的表达水平较低,在预蛹期开始上升;而BmFoxL2-2的表达在5龄幼虫末期达到高峰后开始下降,到了预蛹期后逐渐上升。这些表达变化与二型精子发生的时期相近,推测两个BmFoxL2亚族基因的功能可能与家蚕精子的二型分化相关。
精子发生的过程是一个复杂且精细的过程,在黑腹果蝇中,生殖干细胞通过增殖与分化首先形成性原细胞。一个性原细胞经历有丝分裂和减数分裂后形成64个精细胞,这些过程受到Vasa、Nanos以及细胞周期相关基因的调控(Fabian and Brill, 2012; Fairchildetal., 2016)。研究显示,与黑腹果蝇相似,家蚕生殖干细胞增殖与分化的过程同样需要Vasa和细胞周期等基因的参与(贺真等, 2020)。本研究的结果显示,在Bm12细胞中过表达BmFoxL2-2蛋白可以显著上调细胞周期基因BmCyclinA、BmCyclinB、BmCyclinB3和BmCyclinA,以及生殖干细胞发育关键基因BmNanos和BmVasa的表达(图5)。另外,过表达BmFoxL2-1蛋白也可以上调BmCyclinA和BmNanos的表达,但效果不如BmFoxL2-2的显著(图4)。由此推测家蚕FoxL2亚族蛋白都可以参与精子发生过程。但由于BmFoxL2-1和BmFoxL2-2蛋白的理化性质和亚细胞定位均存在差异(图2),导致了两者对精子发生相关基因的表达调控存在差异。
综上,本研究结果说明BmFoxL2亚族蛋白的功能与精子发生过程相关。以上结果完善了FoxL2亚族基因在雄性生殖系统发育中的功能,同时也为研发通过提高精子质量优化家蚕性状的技术提供理论依据。
猜你喜欢
四川蚕业(2022年2期)2022-11-19
四川蚕业(2022年2期)2022-11-19
现代医学与健康研究电子杂志(2021年21期)2021-12-26
皮肤性病诊疗学杂志(2021年5期)2021-11-27
四川蚕业(2021年2期)2021-03-09
四川蚕业(2021年1期)2021-02-12
三农资讯半月报(2021年1期)2021-01-27
祝您健康·文摘版(2019年1期)2019-05-14
中国医药导报(2016年33期)2017-03-06
癌变·畸变·突变(2015年3期)2015-02-27
