
黑襟毛瓢虫对莲缢管蚜的捕食作用
2022-03-24 08:59杨良波桑海东徐金星唐记平郑兴汶王建国
环境昆虫学报 2022年1期
文 扬,杨良波,桑海东,魏 松,徐金星,唐记平,3,郑兴汶,3*,王建国*
(1. 江西农业大学农学院植保系,南昌 330045;2. 广昌县白莲科学研究所,江西抚州 344900;3. 广昌县白莲产业发展局,江西抚州 344900)
近年来,本课题组在广昌县开展子莲区域性害虫和天敌种类普查,以期摸清害虫及天敌种类数量本底,为探究子莲害虫生物防治提供重要基础。2018年7月份在广昌地区莲田自然生境中首次发现莲缢管蚜周围存在一种类似外形类似介壳虫的昆虫,当时仅取样保存而并未引起重视,在2019年7月份再次发现此昆虫存在,并有一定的种群数量,进而现场仔细观察发现此昆虫对莲缢管蚜有捕食行为,遂引起关注,取样后经昆虫分类学专家虞国跃研究员形态鉴定且分子复核确定该昆虫为黑襟毛瓢虫。
黑襟毛瓢虫Scymnus(Neopullus)hoffmanni属鞘翅目Coleoptera瓢虫科Coccinellidae小毛瓢虫亚科Coccinellidae小毛瓢虫属Scymnus,其主要特点为体型小、繁殖力高、抗逆力强、成虫寿命长(周丽丽, 1986),且具有较好的气候适应性(李文凤等, 2019)。黑襟毛瓢虫的食谱包括棉蚜AphisgossypiiGlover、麦长管蚜SitobionavenaeFabricius、桔蚜ToxopteracitricidusKirkaldy、夹竹桃蚜AphisneriiBoyer、豆蚜AphiscraccivoraKoch、萝卜蚜LipaphiserysimiKaltenbach(杨建武, 1990)。1963年刘崇乐教授对该瓢虫进行了首次报道,此后多地对其分布作了调查,但总的来说,对其研究尚浅,仅仅局限于黑襟毛瓢虫生活习性初步观察、温度对黑襟毛瓢虫发育的影响、黑襟毛瓢虫对棉蚜的数值反应、黑襟毛瓢虫的生物学特性及有效积温及与其它小毛瓢虫的亲缘关系等研究(赵鼎新和王忠文, 1987a; 赵鼎新, 1987b; 杨建武和郑瑞丰, 1990; 黄卫东等, 2019)。黑襟毛瓢虫作为白莲重要害虫莲缢管蚜的天敌昆虫,仅见郑兴汶(2016)等人将黑襟毛瓢虫作为该害虫的天敌昆名录列出(郑兴汶等, 2016),对于其它方面尚无报导。因此,本文以黑襟毛瓢虫为研究对象,通过测定黑襟毛瓢虫各龄幼虫及成虫对莲缢管蚜的捕食功能反应、寻找效应、种内干扰对其捕食能力的影响,旨在揭示黑襟毛瓢虫对莲缢管蚜的捕食潜力,为今后作为莲缢管蚜生物防治天敌昆虫产品的开发和利用提供坚实的理论基础。
1 材料与方法
1.1 供试虫源
莲缢管蚜:于2019年7月上旬采自江西省抚州市广昌县广昌白莲科技博览园苗圃,试验所用蚜虫为2~3龄无翅若蚜。
黑襟毛瓢虫:于2019年7月上旬年采自江西省抚州市广昌县白莲科技博览园苗圃,在人工气候箱内以莲缢管蚜饲养,待成虫产卵孵化后,选取各龄幼虫及成虫,饥饿24 h后供试。
1.2 供试子莲材料
饲养材料选用未被莲缢管蚜为害的健康白莲嫩莲梗,采自江西省抚州市广昌县白莲科技博览园苗圃内,带回室内进行剪切,长度为约5 cm,用浸水的脱脂棉包裹嫩莲梗两端,防止叶片失水同时保持空气湿度。
1.3 试验条件和试验容器
试验在光照培养箱箱内(温度26±1℃、RH70%±5%、光周期16 L ∶8 D,上海博讯医疗生物仪器股份有限公司,BSG-300)进行,采用90 mm×15 mm的培养皿作为试验容器(赵鼎新和王忠文, 1987a)。
1.4 试验方法
1.4.1黑襟毛瓢虫对莲缢管蚜的捕食功能反应
在培养皿放入准备好的白莲嫩茎,并用脱脂棉球保湿,选取2~3龄的健康无翅若蚜,使用毛笔将其挑入培养皿,之后将1头饥饿24 h的黑襟毛瓢虫幼虫或成虫。蚜虫密度设5个处理,每一密度设5次重复,黑襟毛瓢虫幼虫或成虫对莲缢管蚜的捕食密度:1龄幼虫设置2、4、6、8、10头/皿蚜虫,2龄幼虫设置10、15、20、25、30头/皿蚜虫,3龄幼虫设,置20、30、40、50、60头/皿蚜虫,4龄幼虫设置25、35、45、55、65头/皿蚜虫,成虫设置30、40、50、60、70头/皿蚜虫,置于光照培养箱中,24 h后观察记录各培养皿中剩下的和自然死亡的蚜虫数,并测定黑襟毛瓢虫的日捕食量(王倩倩等, 2019)。
1.4.2黑襟毛瓢虫捕食的种内干扰效应
培养皿中接入100头莲缢管蚜,方法同1.4.1,按5个梯度放置黑襟毛瓢虫成虫,分别为每皿1头、2头、3头、4头和5头,然后置于光照培养箱中,24 h后检查记录各培养皿莲缢管蚜剩余数量和自然死亡的蚜虫数量,每个密度5次重复(王倩倩等, 2019)。
1.4.3黑襟毛瓢虫田间捕食观察
采用目测法,仔细观察黑襟毛瓢虫捕食特性,于2019年5-7月间不定期定株(81株)检查莲浮叶、立叶、花、花梗、叶梗等部位黑襟毛瓢虫的卵、幼虫、蛹、成虫等总量(王兴民等, 2014)。
1.5 分析方法
1.5.1捕食功能反应
采用Holling-Ⅱ型圆盘方程进行拟合,公式为:Na=aTN/(1+aThN)(吴坤君等, 2004),式中Na为被捕食的猎物数量;N表示猎物密度;a表示捕食者对猎物的瞬时攻击率,T表示猎物暴露给捕食者的时间(本次试验的时间为1 d),Th为捕食1头猎物所需要花费的时间。
1.5.2寻找效应
如果黑襟毛瓢虫对莲缢管蚜的捕食反应符合Holling-II型功能反应模型,则利用拟合模型所得到的参数估算黑襟毛瓢虫各龄幼虫及成虫对莲缢管蚜的寻找效应S,S=a(1+aThN)(巫鹏翔等, 2017; 胡进锋等, 2017; 喻会平等, 2018; 马宝旭等, 2018)。
1.5.3干扰效应
采用Hassell模型E=QP-m拟合(Hassell, 1969),针对天敌密度对捕食率的影响进行分析。式中Q为P=1头时的最大捕食量;m为干扰系数;E为单头捕食者平均捕食量;P为捕食者密度。
1.5.4数据分析
各模拟方程的理论值与实际值的卡方(χ2)检验均应用SPSS 17.0软件进行统计分析,参数用最小二乘法估计。
2 结果与分析
2.1 黑襟毛瓢虫对莲缢管蚜的捕食功能反应
黑襟毛瓢虫各龄幼虫及成虫的日捕食量随莲缢管蚜密度的增大而增加,当到达一定程度时捕食量会趋于稳定,其功能反应曲线呈负加速曲线,是逆密度制约的。对照Holling提出的3种不同的功能反应函数,符合其中的Holling-Ⅱ型功能反应(Hassell, 1969; Moosa and Zahra, 2013),使用Holling-Ⅱ圆盘方程Na=aTN/(1+aThN)来拟合试验数据,用最小二乘法拟合方程并得到各参数(表1)。其方程的相关系数r=0.9642-0.9860>r(0.01, 3)=0.959,表明黑襟毛瓢虫的捕食量与莲缢管蚜的密度极显著相关。用拟合方程计算得到的理论值与实测值进行卡方检验,χ2=0.2500-1.2727<χ2(0.05, 4)=9.488,黑襟毛瓢虫的捕食量实测值与理论值无显著差异,拟合的Holling-II型圆盘方程能够较好的反映不同莲缢管蚜密度下黑襟毛瓢虫的捕食变化规律。黑襟毛瓢虫幼虫对莲缢管蚜的瞬间攻击率a随着虫龄的增大而增大,黑襟毛瓢虫幼虫1~4龄虫态处理时间Th随之缩短,日最大捕食量1/Th也依次增大,分别为5.5、6.8、14.7和15.1头。成虫的瞬间攻击率大于各个龄期幼虫,处理时间短于各龄幼虫,当N→+∞时,可得单头黑襟毛瓢虫对莲缢管蚜日最大捕食量为22.9头,明显高于各龄幼虫。黑襟毛瓢虫各虫态对莲缢管蚜的捕食能力,即a/Th值,依次为成虫> 4龄幼虫> 3龄幼虫>2龄幼虫>1龄幼虫。

表1 黑襟毛瓢虫对莲缢管蚜的捕食功能反应
2.2 黑襟毛瓢虫对莲缢管蚜的寻找效应
在莲缢管蚜不同密度处理中,黑襟毛瓢虫各龄期幼虫及成虫的寻找效应随着猎物密度的增加而降低(图1)。寻找效应与猎物密度之间的关系呈线性相关,采用最小二乘法进行拟合,黑襟毛瓢虫1~4龄幼虫和成虫的寻找效应与猎物密度间的线性方程分别为y=-0.03x+0.5899 (r=0.9782>r(0.01, 3)=0.9587)、y=-0.0091x+0.4318 (r=0.9667>r(0.01, 3)=0.9587)、y=-0.0026x+1.0022 (r=0.9997>r(0.01, 3)=0.9587)、y=-0.0047x+0.4824 (r=0.9860>r(0.01, 3)=0.9587)、y=-0.0064x+0.7024 (r=0.9679>r(0.01, 3)=0.9587)。黑襟毛瓢虫1龄幼虫的寻找效应随猎物密度增加而下降的趋势最大,3龄幼虫下降的趋势最小。在相同的猎物密度下,3龄幼虫的寻找效应都明显高于其它虫龄的寻找效应。
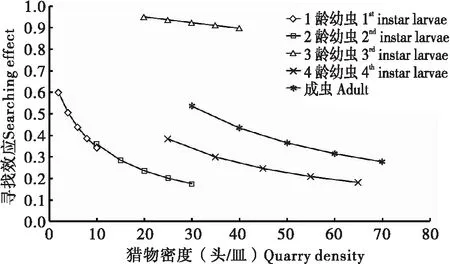
图1 黑襟毛瓢虫寻找效应与莲缢管蚜密度的关系Fig.1 Relationships between searching effects of Scymnus (Neopullus) hoffmanni Weise and the density of Rhopalosiphum nymphaeae (Linnaeus)
2.3 黑襟毛瓢虫捕食的种内干扰效应
当莲缢管蚜的密度为100头/皿时,黑襟毛瓢虫成虫对莲缢管蚜的平均捕食量随着自身密度的增加而逐渐降低,捕食作用率(E)也相应的降低,说明黑襟毛瓢虫成虫在捕食莲缢管蚜时,个体间存在相互干扰和竞争。根据Hassell提出的干扰反应模型(Hassell, 1969):E=Q·P-m,计算出搜索常数Q为0.2202,干扰系数m为0.7220。黑襟毛瓢虫捕食莲缢管蚜过程中的自我干扰方程为:E=0.2202P-0.722。相关系数r=0.9994>r(0.01, 3)=0.9587,表明黑襟毛瓢虫成虫单头的日平均捕食量与其自身密度极显著相关。经卡方检验,得χ2=3.03448<χ2(0.05, 4)=9.488,结果表明,上述模型试验数据与模型的拟合性良好,可以用来描述黑襟毛瓢虫捕食过程中的自干扰。
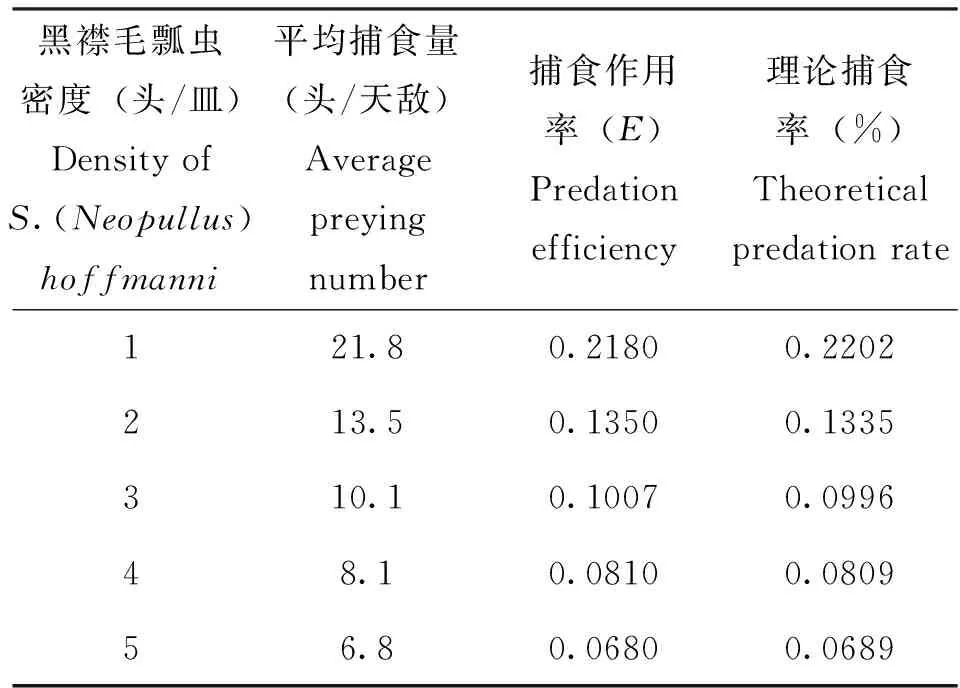
表2 不同密度下黑襟毛瓢虫对捕食量的干扰反应
2.4 黑襟毛瓢虫莲田中的生活及捕食习性观察
黑襟毛瓢虫在田间主要集中在莲植株下部活动,其中莲浮叶是其主要的栖息场所,较其它部位聚集数量更多,81株莲株中最多时可聚集217头(6月28日),其次是荷梗,81株莲株莲梗最多时发现了97头黑襟毛瓢虫。4次调查结果显示,莲的花朵上均未发现黑襟毛瓢虫活动踪迹(图2)。
通过田间观察发现,黑襟毛瓢虫的成虫兼具爬行和飞行能力,可在一定范围的莲株上迁移、活动。幼虫活动范围较小,基本集中在邻近的莲株上活动,其中3龄幼虫爬行速度最快。黑襟毛瓢虫成虫和幼虫均可捕食莲缢管蚜。成虫快速四处寻觅食物,遇到莲缢管蚜立即将其咬住,后用口器咀嚼吞食整头蚜虫。幼虫则在爬行时张开下额须,一旦蚜虫触碰到其下额须便迅速咬住蚜虫的管腹或胸足,再用中、后足固定在莲叶或莲梗上,前足辅助抓握,咬穿蚜虫体壁,昂起头部,左右甩动蚜虫,吸食体液。
3 结论与讨论
根据反应曲线的形状Holling提出将捕食功能反应分为3种基本类型。在本试验条件下,黑襟毛瓢虫对莲缢管蚜的捕食功能反应随着莲缢管蚜密度的增加而增加,其捕食率随着莲缢管蚜的密度呈负增加,这与Holling-Ⅱ型功能反应模型相一致(吴坤君等, 2004)。本试验结果显示,黑襟毛瓢虫对莲缢管蚜有较好的捕食能力,成虫的捕食能力强于幼虫,1~4龄幼虫及成虫对莲缢管蚜的日最大捕食量分别是5.5、6.8、14.7、15.1和22.9头。黑襟毛瓢虫相比较其它瓢虫的捕食量更少,例如异色瓢虫Harmoniaaxyridis成虫对胡萝卜微管蚜Semiaphisheraclei(Takahashi)的理论最大日取食量能够达到83头(李新兵等, 2019),其原因主要由黑襟毛瓢虫体型相对那些瓢虫而言更小,日取食量也相对下降,但黑襟毛瓢虫有其自身优势,例如繁殖力高、抗逆力强、成虫寿命长。天敌对害虫的瞬时攻击率a与处理时间Th的比值是衡量天敌作用效果的参数之一,a/Th值越大表示对害虫的控制能力越强(牟吉元等, 1997; 张伟等, 2018; 喻会平等, 2018; 孙莉等, 2019),本研究中,黑襟毛瓢虫成虫对莲缢管蚜的a/Th值明显大于其各龄幼虫对莲缢管蚜的a/Th值,黑襟毛瓢虫成虫的a/Th达40.90。捕食功能反应可能受多种因素的干扰,例如捕食者的生理状态、空间分布、环境因子等,但本实验结果在一定程度上表明了黑襟毛瓢虫对莲缢管蚜具有较好的防治效果。
黑襟毛瓢虫各龄幼虫及成虫对莲缢管蚜的寻找效应随着猎物密度的增加而降低,这种现象在捕食者与猎物及寄生者与寄主系统中是普遍存在的,例如红彩真猎蝽HarpactorfuscipesFabricius对烟蚜MyzuspersicaeSulzer(邓海滨等, 2015)、加州新小绥螨Neoseiuluscalifornicus对土耳其斯坦叶螨Tetranychusturkestani(汪小东等, 2015)以及十三星瓢虫HippodamiatredecimpunctataLinnzeus对枸杞蚜虫Aphissp.(任月萍, 2015)的寻找效应均随猎物或寄主密度的增加而降低。
黑襟毛瓢虫可能是烟粉虱Bemisiatabaci潜在的捕食天敌候选者之一,尽管目前的研究显示其种群密度较低(Zhangetal., 2007),李文凤等(2019)报道,在云南甘蔗田中,黑襟毛瓢虫等天敌昆虫整年在田间捕食粉虱,能把粉虱危害控制在经济损失水平以下(李文凤等, 2019)。本文结果显示江西广昌实验室和野外条件下,黑襟毛瓢虫对子莲上的莲缢管蚜均有良好的控制效果。
种内干扰对黑襟毛瓢虫的捕食量有较大影响,在蚜虫密度和捕食空间一定的情况下,随着黑襟毛瓢虫群体密度的增加,单头平均捕食量呈下降趋势。二者间关系可以用模型E=0.2202P-0.722拟合。因此,如果需要利用黑襟毛瓢虫防控莲缢管蚜,必须注意保护利用田间自然种群,根据田间天敌和猎物密度合理进行人工释放,从而高效的利用天敌。在田间利用黑襟毛瓢虫防治莲缢管蚜,今后还需要研究该瓢虫越冬等人工扩繁技术、春季田间种群密度和蚜虫密度关系,解决天敌滞后效应及杀虫剂使用与瓢虫生物防治协调问题。
猜你喜欢
小天使·一年级语数英综合(2022年5期)2022-05-25
发明与创新·小学生(2021年5期)2021-08-23
发明与创新(2021年19期)2021-05-07
红蜻蜓·低年级(2020年9期)2020-09-26
落叶果树(2020年2期)2020-04-20
小学生导刊(2018年31期)2018-12-06
小学生作文(低年级适用)(2018年4期)2018-11-29
小朋友·快乐手工(2015年10期)2015-11-02
中国新闻周刊(2015年31期)2015-08-28
小朋友·快乐手工(2015年2期)2015-03-13
