
短链脂肪酸调控奶牛乳腺乳脂合成作用机制的研究进展
2022-03-30 05:20陈美庆张养东王加启
动物营养学报 2022年3期
陈美庆 张养东 郑 楠 王加启
(中国农业科学院北京畜牧兽医研究所,农业农村部奶及奶制品质量安全控制重点实验室,农业农村部奶产品质量安全风险评估实验室,农业农村部奶及奶制品质量监督检验测试中心,动物营养学国家重点实验室,北京100193)
乳脂是牛奶中重要的营养物质,对于牛奶消费者的营养和健康有着积极影响[1]。此外,乳脂是牛奶主要的能量物质,与牛奶生产者的经济利益密切相关。因此,深入了解乳腺中乳脂合成的调节机制对于改善泌乳期间反刍动物的能量平衡和提高牛奶对消费者的营养价值至关重要。短链脂肪酸(short-chain fatty acids,SCFA)是碳原子数不大于6的饱和脂肪酸,因其具有挥发性,常常被称为挥发性脂肪酸[2]。奶牛体内的绝大部分SCFA经瘤胃发酵碳水化合物产生,少部分由肠道微生物发酵产生,多以离子的形式存在,由瘤胃上皮或肠道上皮吸收进入不同组织,参与能量代谢和营养物质代谢等机体内多项生理活动[3]。研究发现,乙酸、丙酸和丁酸是奶牛瘤胃发酵碳水化合物产生的主要代谢产物,能够满足反刍动物60%~80%的能量需求[4]。其中,丙酸是葡萄糖的主要前体物质,以葡萄糖的形式参与机体能量代谢,而乙酸和丁酸一方面作为乳腺内脂肪酸从头合成的前体物,另一方面作为信号分子调节乳腺内脂肪酸代谢,影响奶牛乳脂的组成和含量[5]。本文就SCFA对乳脂合成的影响及其调控乳腺乳脂合成的分子机制2方面,综述了SCFA调控奶牛乳腺乳脂合成的作用机制,为奶牛乳脂合成机制的相关研究提供理论依据。
1 SCFA对奶牛乳脂合成的影响
Urrutia等[6]在奶牛饲粮中添加乙酸盐和丁酸盐发现,添加2.9%乙酸盐显著提高了奶牛乳脂率和乳脂产量,分别提高了0.2%和90 g/d,而在饲粮中添加等碳当量的丁酸盐对乳脂率和乳脂产量无显著影响。Matamoros等[7]在奶牛饲粮中添加3.25%乙酸盐显著提高了奶牛乳脂产量,这可能是因为饲粮中添加乙酸盐增加了乳腺的乙酸盐供应,从而刺激脂肪酸从头合成的产生来增加乳脂产量。Izumi等[8]研究发现,在奶牛饲粮中添加1.1%的丁酸能增加乳脂产量,在一定程度上缓解高精料饲粮引起的奶牛乳脂抑制。Seymour等[9]通过综述奶牛瘤胃SCFA含量与乳成分的关系,发现瘤胃内丁酸含量与产奶量呈正相关,瘤胃内乙酸/丙酸与乳脂产量呈正相关。研究学者们在荷斯坦奶牛的瘤胃灌注SCFA试验时发现,灌注乙酸盐和丁酸盐能显著提高奶牛的乳脂率、乳脂产量和乳中从头合成脂肪酸(<16碳和16碳)含量,其中棕榈酸的含量和产量增加较多,且乙酸盐灌注量与乳脂合成之间存在明显的剂量-效应关系,奶牛乳脂率和乳脂产量随乙酸盐的线性供应分别呈现线性和二次曲线增加;此外,还发现灌注乙酸盐能显著增加瘤胃中乙酸和血浆中β-羟基丁酸(β-hydroxybutyrate,BHB)的含量,对瘤胃中丁酸的含量无显著影响,灌注丁酸盐能显著增加瘤胃中丁酸和血浆中BHB的含量,这表明增加泌乳奶牛的瘤胃乙酸盐和丁酸盐的供应可能通过增加乙酸和BHB的含量,为脂肪酸从头合成提供更多底物进而提高乳脂产量[10-12]。灌注丙酸对奶牛的乳脂含量和产量无显著影响[13]。向荷斯坦奶牛的真胃输注乙酸增加了乳脂产量,这表明乳腺吸收的更多的乙酸盐被用于乳脂合成[14]。
综上所述,乙酸、丁酸以及BHB能增加奶牛乳脂率和乳脂产量,而丙酸对乳脂合成无促进作用。然而,不同SCFA对奶牛乳脂产量和含量的影响程度不同,这可能与具体SCFA在乳腺乳脂合成过程中所发挥的作用相关。
2 SCFA影响奶牛乳腺乳脂合成的作用机制
2.1 作为乳脂合成前体物参与乳腺内脂肪酸合成
乳脂主要由约95%的甘油三酯(TG)、约2%的甘油二酯、约1%的单酰甘油、约1%的磷脂、少于0.5%的胆固醇、少于0.5%的游离脂肪酸组成[15-16]。反刍动物合成乳脂所利用的脂肪酸来源于乳腺上皮细胞的从头合成和外源摄取2个部分。中短链脂肪酸(<16碳)和约50%的16碳脂肪酸约占合成乳脂所利用的脂肪酸的1/2,这部分脂肪酸由乳腺上皮细胞利用乙酸和BHB从头合成,乳脂中16碳以上长链脂肪酸和约50%的16碳脂肪酸经乳腺上皮细胞直接从血液中吸收得到[17-18]。因此,乙酸和BHB是反刍动物乳腺上皮细胞从头合成脂肪酸的前体物质,在乳脂合成中起到十分重要的作用[19]。
乙酸是乳中脂肪酸从头合成的主要碳源,主要通过瘤胃发酵碳水化合物产生,BHB主要通过瘤胃上皮细胞吸收瘤胃发酵产物丁酸产生[20]。这些SCFA被瘤胃上皮细胞或肠上皮细胞吸收进入血液,经血液循环后,通过扩散作用穿过毛细血管内皮和间质空隙进入乳腺上皮细胞利用。奶牛体内的同位素标记试验表明,丁酸提供了从头合成脂肪酸中前4个碳约50%的碳源,而乙酸提供了其余所需的绝大多数碳源[21]。奶牛乳腺上皮细胞内脂肪酸从头合成,涉及脂肪酸的活化、碳链延长、去饱和等过程,此过程需要多种酶的参与[22-24]。乙酸和BHB在乳腺上皮细胞中辅酶A短链家族成员(acetyl coenzyme A synthetase,ACSS)的催化下,激活形成乙酰辅酶A和β-羟基丁酰辅酶A[25]。2分子乙酰辅酶A在乙酰辅酶A羧化酶(acetyl coenzyme A carboxylase,ACACA)催化下羧合形成丙二酰辅酶A,在脂肪酸合成酶(fatty acid synthetase,FAS)的进一步催化下与乙酰辅酶A(或少量β-羟基丁酰辅酶A)依次结合,逐渐连接成4~16碳等不同碳链长度的饱和脂肪酸,以合成16碳饱和脂肪酸为主[26-29]。乳腺从头合成形成的14~16碳饱和脂肪酸在硬脂酰辅酶A去饱和酶(stearoyl coenzyme A desaturase,SCD)的催化下,Δ9位置形成顺式双键,转变为单不饱和脂肪酸[30-31]。
2.2 作为信号分子调控乳腺脂质合成
乙酸、丁酸以及BHB通过影响脂质合成相关信号通路、调控与乳脂合成相关的转录因子和酶的基因表达,调控乳腺脂质合成。
2.2.1 调控乳脂合成关键酶的活性
ACACA催化乙酰辅酶A生成丙二酰辅酶A,是脂肪酸从头合成的限速酶,FAS催化丙二酰辅酶A和乙酰辅酶A合成4~16碳等不同碳链长度的饱和脂肪酸,在乳腺细胞从头合成上起着重要作用,SCD1是单不饱和脂肪酸合成的限速酶。在乳脂合成过程中,ACACA、FAS、SCD1是乳脂合成的限速酶,其活性大小反映了乳腺乳脂合成作用的程度,关键酶的mRNA表达水平可反映相应的酶活性。因此,通过提高乳脂合成关键酶的mRNA表达水平可以促进乳脂合成[32]。
研究表明,乙酸能显著提高奶牛乳腺上皮细胞(bovine mammary epithelial cells,BMECs)内TG的含量,上调FAS和ACACA的mRNA表达水平[33],显著提高奶牛永生乳腺上皮细胞(bovine mammary cell line,MAC-T)内TG的含量,上调ACACA和SCD1的mRNA表达水平[34]。研究表明,乙酸能通过上调过氧化物酶增殖物激活受体γ(peroxisome proliferator-activated receptor,PPARγ)和固醇调控元件结合蛋白1(sterol regulatory element-binding protein 1,SREBP1)的mRNA表达水平,增加脂肪酸合成相关酶ACACA、FAS和SCD1的mRNA和蛋白表达水平,增加脂肪酸的从头合成和去饱和,从而促进乳脂合成[35-36]。Cheng等[37]研究发现,在BMECs中丁酸钠提高了FAS和ACACA的mRNA表达水平,促进乳脂合成。Zhang等[38]研究发现,BHB能显著增加FAS和ACACA的mRNA和蛋白表达水平,从而促进乳脂合成。Ali等[39]在BMECs中添加不同浓度的单独及组合的乙酸盐和BHB,发现乙酸盐和BHB以剂量依赖性方式增加乳腺细胞内TG含量和脂滴形成,且乙酸盐和BHB及其联合作用使ACACA、FAS和SCD1的mRNA表达水平和SREBP1的蛋白表达水平与TG含量和脂滴形成的变化一致,进一步说明乙酸盐和BHB可能通过激活BMECs中的SREBP1信号通路作用于其调节乳脂合成的靶向基因。不同的是,Jacobs等[34]在MAC-T中分别添加乙酸盐和BHB,发现乙酸盐能促进ACACA和SCD1的mRNA表达水平,但不影响SREBP1的mRNA表达水平,而BHB对ACACA和SCD1的mRNA表达水平无显著影响。
2.2.2 调控乳脂合成相关转录因子的基因表达
SREBP1是碱性螺旋环亮氨酸拉链家族的一员,是脂肪合成的主要转录调节因子[40]。PPARγ属于核受体超家族成员,可作为细胞内脂肪酸的生物传感器,改变脂质代谢[41]。PPARγ和SREBP1是调控乳脂合成的关键转录因子,可通过调控其下游的靶基因ACACA、FAS和SCD1等参与脂肪酸从头合成、脂肪酸去饱和等过程,进而调控乳脂代谢[25, 35, 40]。
Liu等[35]通过PPARγ基因沉默技术改变BMECs中PPARγ的表达,并同时用12 mmol/L乙酸钠处理BMECs后发现,对BMECs中的PPARγ进行基因沉默显著减低了SREBP1、ACACA、FAS和SCD的mRNA和蛋白表达水平,显著降低了细胞活力与细胞增殖,显著降低了BMECs培养液中TG的含量;而添加乙酸钠显著提高了PPARγ基因沉默的BMECs中PPARγ和SREBP1的蛋白表达水平以及PPARγ、SREBP1、ACACA、FAS和SCD的mRNA表达水平,显著增加了BMECs培养液中TG的含量,因此,乙酸可通过PPARγ在乳腺细胞生长、脂肪酸从头合成、脂肪酸去饱和、TG合成等过程发挥重要作用。Zhang等[38]通过SREBP1基因沉默技术改变BMECs中SREBP1的表达,并同时用BHB处理BMECs后发现,对BMECs中的SREBP1进行基因沉默显著减低了SREBP1、ACACA和FAS的mRNA和蛋白表达水平,显著降低了BMECs培养液中TG的含量;而添加BHB显著增加了SREBP1基因沉默的BMECs中SREBP1的核易位,显著提高了SREBP1的蛋白表达水平以及ACACA和FAS的mRNA表达水平,显著增加了BMECs培养液中TG的含量,因此,BHB可通过SREBP1在乳腺细胞生长、脂肪酸从头合成、脂肪酸去饱和、TG合成等过程发挥重要作用。Ali等[39]在BMECs中添加不同浓度的单独及组合的乙酸盐和BHB,发现乙酸盐和BHB以剂量依赖性方式增加乳腺细胞内TG含量和脂滴形成,且乙酸盐和BHB及其联合作用使ACACA、FAS和SCD1的mRNA表达水平和SREBP1的蛋白表达水平与TG含量和脂滴形成的变化一致,进一步说明乙酸盐和BHB可能通过激活BMECs中的SREBP1信号通路作用于其调节乳脂合成的靶向基因。
2.2.3 调控乳脂合成信号通路
哺乳动物雷帕霉素靶蛋白(mammalian target of rapamycin,mTOR)作为丝氨酸/苏氨酸蛋白激酶,在翻译水平上通过影响mRNA与真核细胞翻译起始因子4E(eIF4E)的结合以及真核细胞翻译起始因子4E结合蛋白1(4EBP1)、核糖体S6激酶(S6K)的磷酸化,调控细胞生长、蛋白质合成和脂质代谢[42-44]。其中,SREBP1和PPARγ等脂质合成基因表达相关转录因子在调节乳脂合成的mTOR信号通路中起重要作用[42, 45-46]。激活mTOR信号通路能通过eIF4E、4EBP1和S6K等主要下游效应因子,上调SREBP1和PPARγ等转录调控因子的表达,促进乳脂合成[47]。Zhao等[36]研究发现,乙酸能通过mTOR/eIF4E信号通路,上调SREBP1的表达,增加ACACA、FAS和SCD1的mRNA和蛋白表达水平,以剂量依赖的方式促进BMECs中的乳脂合成,其中4~6 mmol/L乙酸盐的BMECs中mTOR的mRNA表达水平增加了20%~68%,10~12 mmol/L乙酸盐的BMECs中mTOR的mRNA表达水平减少了7%~14%。
腺苷酸蛋白激酶(activated protein kinase-activated protein kinase,AMPK)是一种细胞能量传感器,在乳腺营养代谢和乳成分合成等方面具有重要作用[48-49]。激活的AMPK会抑制MAC-T中乳脂合成[50]。G蛋白偶联受体(G-protein-coupled receptors,GPCR)作为SCFA的受体,接受SCFA的刺激,将信号转导至细胞内,以调控机体代谢。G蛋白偶联受体41(GPR41)、G蛋白偶联受体43(GPR43)和G蛋白偶联受体109A(GPR109A)是目前研究的SCFA的主要受体形式,在不同细胞中介导SCFA的受体各不相同,在BMECs中主要通过GPR41介导SCFA信号[51]。研究发现,乙酸和丁酸通过激活GPR41促进BMECs中AMPK的磷酸化,抑制乳腺内的乳脂合成[37, 49]。mTOR和NAD依赖性脱乙酰酶(SIRT1)作为调节脂质代谢的关键因子,位于AMPK下游,受AMPK调节影响乳脂合成[52-53]。Cheng等[37]研究发现,在BMECs中丁酸钠通过激活GPR41及其下游信号通路,增加SREBP1的核转位和乙酰化,提高FAS和ACC的mRNA表达水平,促进乳脂合成。丁酸钠调控BMECs乳脂合成通过以下2条信号通路,一是GPR41/AMPK/mTOR/S6K/SREBP1信号通路,即丁酸钠通过激活GPR41抑制磷酸化腺苷酸蛋白激酶(p-AMPK)和SIRT1蛋白表达,促进成熟SREBP1的乙酰化;二是GPR41/AMPK/SIRT1/SREBP1信号通路,即通过激活GPR41抑制p-AMPK蛋白表达,上调磷酸化哺乳动物雷帕霉素靶蛋白(p-mTOR)/磷酸化核糖体S6激酶(p-S6K)和成熟SREBP1蛋白表达,提高FAS和ACC的mRNA表达水平,促进乳脂合成。本文归纳了SCFA参与乳腺脂肪酸代谢的相关途径见图1。
综上所述,SCFA可能通过激活GPCR来调节BMECs中AMPK的磷酸化,进而调控ACACA的磷酸化或去磷酸化来控制脂肪生成和脂质氧化,或调控控制SREBP1和PPARγ等脂质合成转录调控因子等上游靶标mTOR的活性,进而调控乳腺内脂质合成。
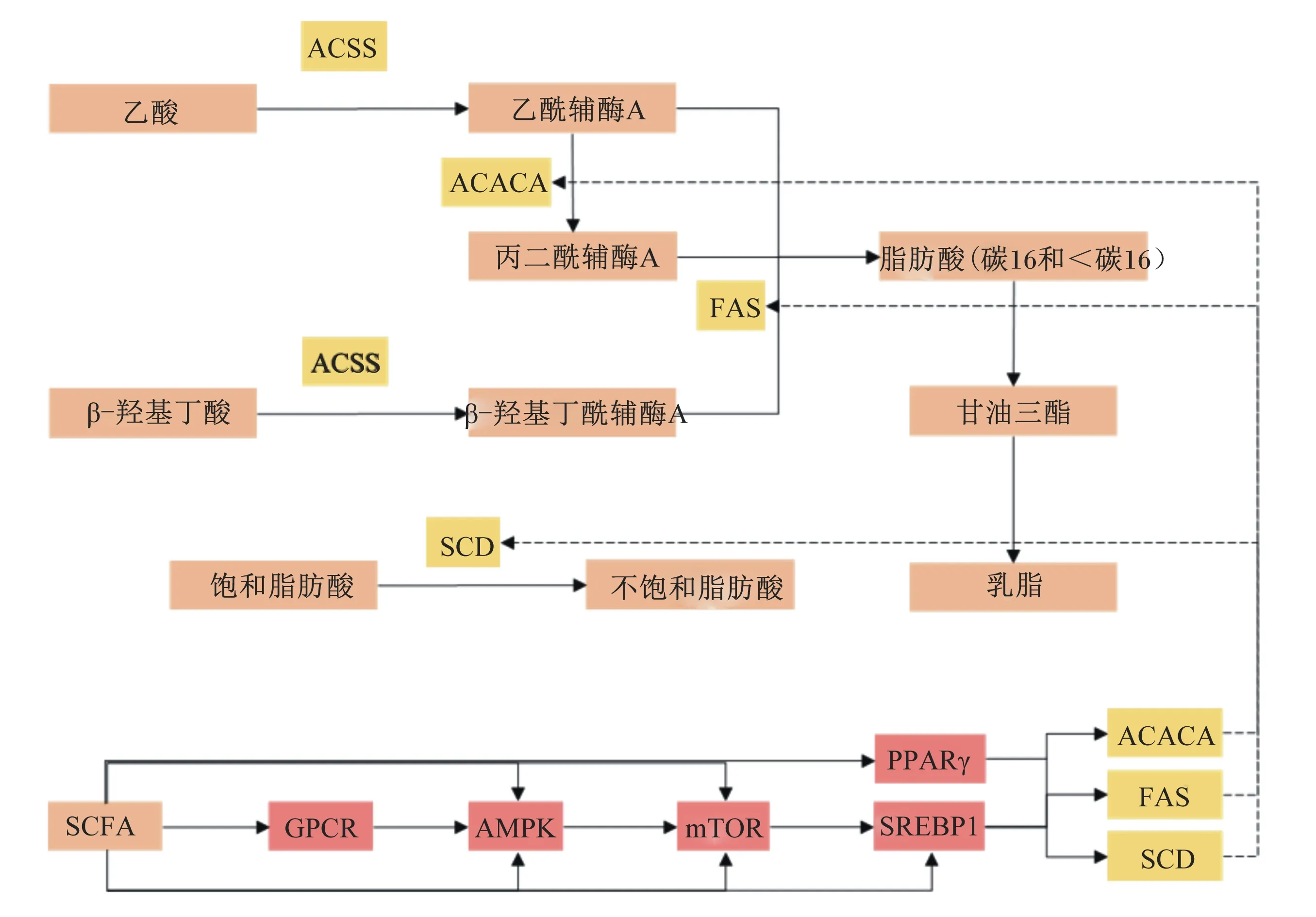
ACSS:辅酶A短链家族成员 acetyl coenzyme A synthetase;ACACA:乙酰辅酶A羧化酶 acetyl coenzyme A carboxylase;FAS:脂肪酸合成酶 fatty acid synthetase;SCD:硬脂酰辅酶A去饱和酶 stearoyl coenzyme A desaturase;SCFA:短链脂肪酸 short-chain fatty acid;GPCR:G蛋白偶联受体 G-protein-coupled receptors;AMPK:腺苷酸蛋白激酶 activated protein kinase-activated protein kinase;mTOR:哺乳动物雷帕霉素靶蛋白 mammalian target of rapamycin;PPARγ:过氧化物酶增殖物激活受体γ peroxisome proliferator-activated receptor γ;SREBP1:固醇调节元件结合蛋白1 sterol regulatory element-binding protein 1。
3 小 结
乙酸、丁酸以及BHB能增加奶牛乳脂率和乳脂产量,而丙酸对乳脂合成无促进作用。其中,乙酸、丁酸以及BHB对乳腺乳脂合成的调控作用主要包括2方面,一是作为乳腺内脂肪酸从头合成的前体物,直接参与乳脂合成;二是作为信号分子,调控乳腺脂质合成。乙酸、丁酸以及BHB通过影响脂质合成相关信号通路、调控与乳脂合成相关的转录因子和酶的基因表达,调控乳腺脂质合成。SCFA可以通过激活GPR41调控AMPK和mTOR等信号通路的活性,调控控制SREBP1和PPARγ等转录调控因子的基因表达,调节乳脂合成相关酶的活性,进而影响BMECs的乳脂合成,但SCFA是通过影响哪些酶、基因、转录因子、信号通路进而影响乳脂合成还缺乏全面清晰认识。在调控机制上,目前对SCFA调控乳腺乳脂合成的研究集中在细胞水平的验证,鲜有报道在奶牛个体水平上SCFA调控乳脂合成的机制。因此,有必要借助基因组学、转录组学、表观遗传组学、蛋白质组学、代谢组学和脂质组学等技术手段分析乳脂合成相关酶、基因、转录因子和信号通路,进一步研究SCFA对乳脂合成的调控机制,这对充分发挥奶牛泌乳潜力、生产优质牛奶具有重要科学意义。
猜你喜欢
动物营养学报(2022年9期)2022-10-12
中老年保健(2022年4期)2022-08-22
当代水产(2022年1期)2022-04-26
当代水产(2022年2期)2022-04-26
中国饲料(2021年17期)2021-11-02
当代水产(2021年2期)2021-03-29
华声文萃(2019年4期)2019-09-10
文萃报·周二版(2019年10期)2019-09-10
分析化学(2018年4期)2018-11-02
现代畜牧科技(2018年3期)2018-05-14
