
基于布尔网络的造血细胞谱系分化选择分析
2022-04-14 14:07王瑞琦陈庆曦叶坚挺
河南师范大学学报(自然科学版) 2022年3期
王瑞琦,陈庆曦,叶坚挺
(上海大学 理学院,上海 200444)
1 问题描述
造血细胞谱系中所有细胞类型的数量通过造血干细胞(Hematopoietic Stem Cells,HSCs)不断生长分化的方式保持动态平衡[1-2].若体内不同细胞类型之间的数量平衡因外部因素影响而被破坏,则会引发严重的血液疾病,如白血病等[3].为了解此类疾病的发病原因以及寻找治疗方法,需要了解正常条件下影响造血细胞分化选择的因素.
通常情况下,生物实验的结果最为可靠且直观,但进行广泛试验意味着较高的试验成本与实验周期,因此引入数学建模的分析方式.布尔网络作为用于定性分析的建模方式,广泛应用于构建基因调控网络,也同样应用于造血细胞谱系分化的分析中[4-8].在布尔网络中可以利用基因之间的调控作用获得调控规则,进而模拟细胞内基因状态随时间的变化过程以及最终稳态.
布尔网络在用于模拟造血细胞谱系分化时,通常针对部分细胞类型进行建模,这意味着会忽略一些重要的细胞转化过程.而增加网络涉及的细胞类型时,网络构建过程所需的节点和调控关系也同步增加,使得构建更为全面的网络时工作量更多,难度也更大.因此考虑通过整合现有小网络的方式在网络全面性和建模工作量之间取得平衡.
2 建模过程
2.1 确定分析范围
为了更加全面地分析造血细胞的分化过程,从造血细胞谱系的经典分化路径中选择了几种主要的细胞类型[2].通常来说,HSCs被看作是造血细胞分化的起点,但在HSCs分化到本文主要讨论的分化阶段之前还需要经历一系列的生长分化过程,因此本文以多能祖细胞(Multipotent Progenitors,MPs)作为分化的起始细胞类型,具体的分化路径如图1所示.MPs向不同的细胞类型分化的选择过程主要由不同的转录因子影响[9].为了构建一个更加全面的涉及更多细胞类型的基因调控网络,利用了两个已知的有关造血细胞谱系分化的基因调控网络[7-8].两个网络都只讨论了与造血细胞谱系中的部分细胞类型有关的转录因子.因此,本文通过特定的整合规则将两个子网络进行整合,从而获得一个可以同时讨论包括巨噬细胞、B细胞、粒细胞和红细胞在内的不同细胞类型间的调控网络,使得对于造血细胞谱系分化过程有更加全面的理解.获得的整合网络不但能够保持原有子网络所能够反映的部分细胞类型的调控规则,同时可以利用两个子网络之间的联系获得更多细胞类型之间的转化关系.因此本文的首要任务是通过特定的整合规则将两个子网络进行整合,同时通过广泛的文献查找获得额外的基因调控规则,对整合后的网络加以修正.

2.2 构建整合调控网络
在整合两个网络时,以两个子网络中共有的4个节点(Cebpa,Spi1,Gfi1和Egr2)作为连接点,使得这4个节点同时在两个网络中生效,并将分属于两个网络中的节点利用调控规则连接到这4个公共节点上,从而获得一个结构整合后的调控网络.除了结构整合之外,还应当把两个网络中公共节点对应的布尔调控规则进行整合.在整合的过程中,由于公共节点在两个子网络中均存在相应的调控规则,因此对两条调控规则的整合方式进行了规定.假如两条规则所涉及的布尔规则完全不同,将其理解为两条规则在实际调控作用中需要同时生效,因此通常利用“&”语句,也就是“和”逻辑进行整合,表示两条规则在影响相应节点表达情况时需要同时生效.假如两条规则中的内容有部分相似,则利用“|”语句,也就是“或”逻辑进行整合,表示这两条规则可以单独发挥作用.另外在整合时,为了网络的精简考虑,不区分基因与其对应生成的蛋白质,只采用基因作为网络的节点.通过以上的整合规定,可以获得一个初步的整合网络及其对应的布尔调控规则.
在修正初始整合网络之前,必须确定各个细胞分化阶段的稳态,也就是对应细胞类型的布尔值,从而为判断构建的网络是否准确提供原始依据.因此首先需要补全所有细胞类型基因表达情况的布尔值.对于某一基因来说,可以获得其在部分细胞类型中已有的布尔值以及文献中参考的实验数据[7-8].在所有已知该基因布尔值的细胞类型中,可以在所有布尔值表达为“0”的细胞中确定一个最大的基因表达数值,这确定了令布尔值表达为“0”的阈值,对于所有低于该值的基因表达数值,均可将布尔值定为“0”.这一阈值确定的原则适用于大部分基因.同理可以获得令基因布尔值表示为“1”的最小基因表达数据,也就是另一个阈值,对于所有高于该阈值的实验数据,均可将布尔值定为“1”.因此对于缺少的布尔值,即可通过实验数据与两个阈值进行比较,当低于第一个阈值时,缺少的布尔值表示为“0”,表示这个基因没有被激活,若高于第二个阈值,则布尔值为“1”,表示这个基因已经被激活了.对于未能依靠阈值判定的布尔值,依据实验数据与阈值的距离确定归属.总的来说,结合文献[7-8]中已有的布尔值以及通过阈值判断得到的缺失布尔值,得到了所有细胞类型完整的标准布尔值,如表1所示,表中加黑字体表示的布尔值代表根据阈值判断得到的布尔值,其他表示的布尔值代表参考子网络中原有的布尔值.
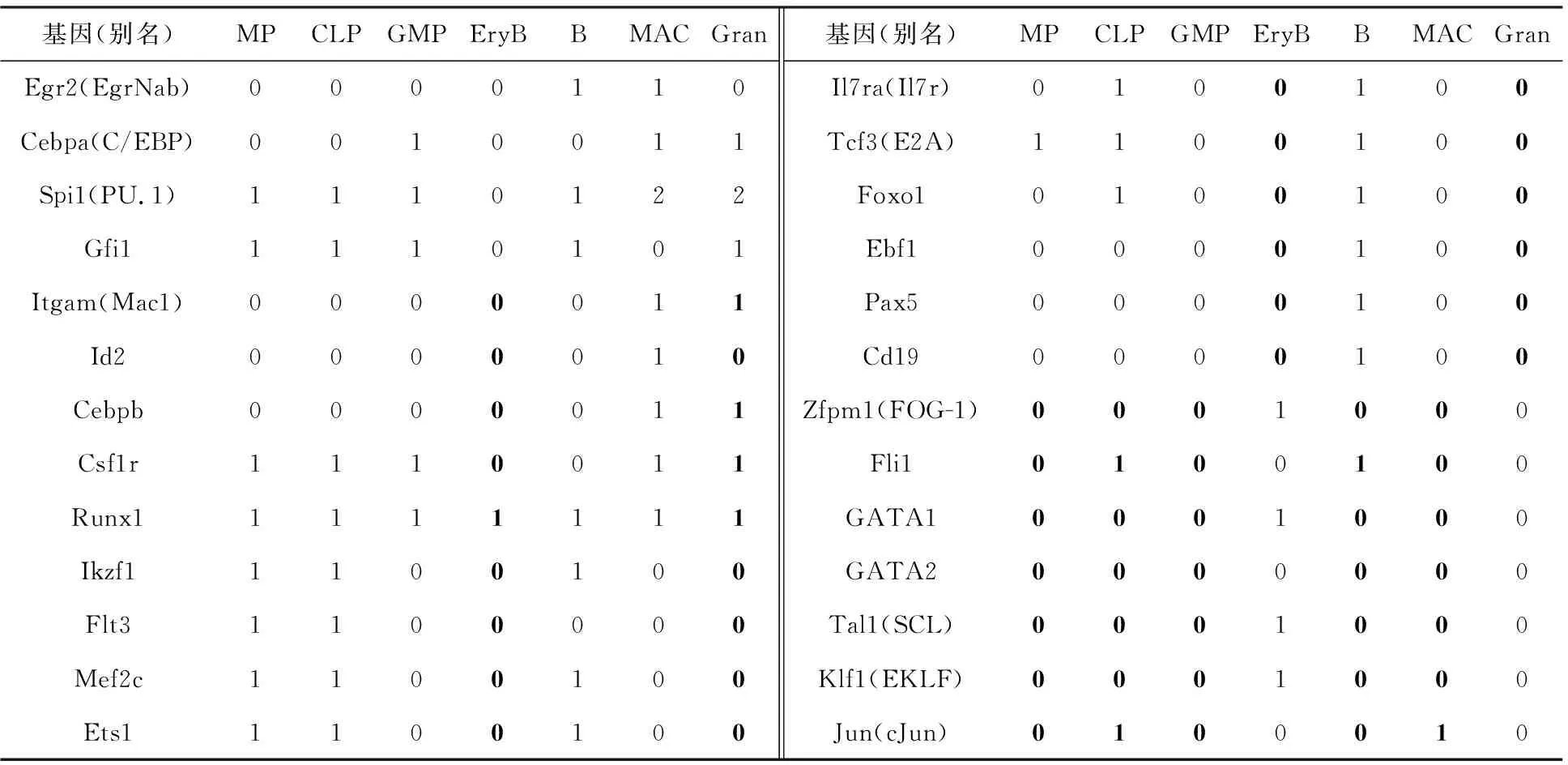
表1 所有细胞类型的布尔值
本文的目的是获得能够反映造血细胞分化情况的整合调控网络,验证网络准确性的方式是判断整合调控网络获得的稳态结果与本文依据阈值和基因表达数据获得的标准布尔值是否匹配,因此需要对整合调控网络的稳态进行分析.利用GINsim运行初步整合调控网络得到布尔值,并与此前确定的标准布尔值进行比较.由于两者的表达情况有明显的差异,通过查阅文献获得了部分实验结果并在现有网络的基础上增加了相应的调控规则[10-18],结果如表2所示.

表2 整合调控网络的布尔调控规则
比较了整合调控网络在修正后所得到的所有细胞的布尔值与文献[19]中的细胞实验表达值.当连续的实验数据与离散的布尔值整体相同时,则说明整合网络可以得到对应细胞类型的稳态结果,比较结果如图2所示.在选择适当的阈值之后,可以发现除GATA-2外的大部分基因在不同细胞类型上的实验表达情况与整合调控网络输出的布尔值类似.对于GATA-2的差别,首先根据文献[8]中子网络的结构,当GATA2被激活之后,由GATA1和Zfpm1所组成的抑制性相干前馈环会重新抑制GATA2,这意味着GATA2在表达时通常是处于持续波动的状态,在稳态中的表现则通常为不表达.同时在文献[8]分析造血细胞分化过程中,当早期祖细胞分化为髓系祖细胞时,GATA2已经被下调,这说明GATA2在GMP等髓系细胞中不会被激活,而根据基因表达数据来看,GATA2在GMP中的表达值为最高,这意味着GATA2在所有细胞中都不会被激活,另外由于抑制性相干前馈环的作用,GATA2的变化过程会被延迟,而GATA2的实验表达数据在GMP细胞向MAC细胞转化过程中显著减少说明GATA-2在分化过程中确实会明显下降,同样说明GATA2在分化潜能较低的这几类细胞类型中不会被激活.总的来说,以上的比较结果意味着构建的整合网络可以再现细胞分化过程中各个细胞类型的基因表达特征,为进一步分析不同细胞类型间的分化作用提供了有效的分析基础.

不同细胞类型的布尔值在整体上与实验表达数据相近,其中GATA-2的差别由抑制性相干前馈环等原因解释.这意味着构建的整合调控网络可以正确地重现细胞分化过程中各细胞类型的基因表达特征.
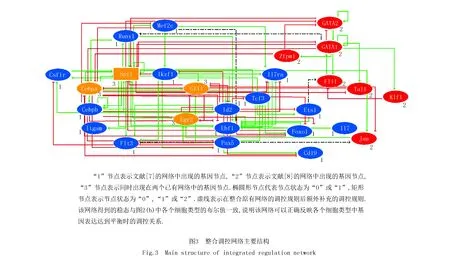
总之,通过结合已有的两个布尔调控网络获得了一个初步的整合调控网络,并利用已知的实验结果对该网络进行修正.最终根据整合调控网络得到的布尔值与实验数据的对比验证了所获整合网络的可靠性,网络主体如图3所示.这一整合调控网络包含了多能祖细胞向下分化所得到的主要细胞类型,因此计划通过施加扰动的方式了解实现细胞定向分化的主要影响因素.
2.3 获得定向分化组合扰动策略
在分化过程中细胞的基因表达情况会发生变化,这些发生变化的基因通常与对应的分化选择有关,但其中只有部分基因起到主导作用,其余的基因则是在前者的诱导下发生变化.当想要将一种类型的细胞定向分化为另一种细胞时,最直接的策略是同时修改分化前后细胞类型之间全部表达有差异的基因,但更为合理的策略是仅扰动起到主导作用的基因.
通过计算机模拟各类扰动策略为验证生物系统的合理性提供了可能,在模拟过程中所有网络中的节点和边都可以作为扰动的对象.对于调控所采取的策略都有实际的实验原理与之对应,例如调控节点的敲除可以通过破坏其调控启动子序列来实现,另一方面,过表达可以通过诱导结构等异位转录因子表达的方式来实现,而调控相互作用的敲除则对应着转录因子与其靶基因的启动子区域的结合亲和力突变[20-23].通过修改布尔模型中的节点状态来实现两种系统性扰动,即瞬态扰动或持续性扰动.对于瞬态扰动,仅在模拟初始时修改被扰动节点的布尔值,使得该节点提供扰动信号给其他后续参与到分化过程的节点,而在模拟过程中,该节点仍可能由于受到其他节点的反馈影响而改变布尔值,这一扰动方式主要是为了模拟实际情况下细胞在受到转录因子瞬时诱导之后发生的分化过程.对于持续性扰动,则通过敲除或过表达等方式保持被扰动节点的布尔值被永久改变,这一方式在实验过程中更容易实现.为了分析各个基因在扰动实验中对于分化选择的影响,测试了7种血细胞类型之间的状态转换过程.
通过系统的组合扰动分析,确定了实现定向分化的最小组合扰动策略(最多扰动3个节点),如表3所示.表中上标“T”或“P”分别表示扰动是瞬时的或持久的.例如,从MP到GMP的转换中的扰动意味着Cebpa在MP处的布尔值为0,通过瞬时扰动转化为1.与扰动策略相关的实验依据如表3所示.这些扰动策略可以为造血细胞谱系规范提供一些实验建议.例如,对于红细胞祖细胞的分化来说,调节红细胞生成的两种最主要的细胞因子是促红细胞生成素(Epo)和干细胞因子(SCF)[24].发现当Spi1的布尔值从“1”扰动为“0”,同时GATA1的布尔值从“0”扰动为“1”,即瞬时或持续性采用“Spi1-0,GATA1-1”这一扰动策略时,可以实现MP向EryB的定向分化.通过查阅文献可知EpoR,也就是促红细胞生成素(Epo)的受体,是GATA1的目标基因[25],而Epo是红细胞体外形成的重要细胞因子[24].同时,对于SCF(c-kit配体)来说,其受体c-kit主要受SCL复合物的影响[26],合成SCL的Tal1的表达被Spi1抑制,被GATA1促进[27].这说明了本文的扰动策略确实与对应的细胞分化过程有关,这些实验结果部分证明了所确定的组合扰动策略在生物学上是合理的.对于两种类型的扰动,即瞬时扰动和持续扰动,得到了相同的结果,这表明扰动策略中确定的节点在造血细胞分化选择上起着重要作用.

表3 实现定向分化的最简扰动策略
3 结 论
本文通过整合两个已有的子网络,构建了一个更全面的基因调控网络来分析在单扰动或组合扰动下如何实现不同类型血细胞之间的定向分化.当整合两个子网时,除了合并两个子网络中原有的调控规则外,还需要补充一些额外的调控规则,使得整合调控网络的逻辑模型所获得的稳态与实验数据保持一致,这样的一致性说明了本文构成的整合调控网络和逻辑模型是合理的.然后,通过确定最小扰动策略,也就是要扰动的最小节点集以及确定如何扰动它们,从而可以实现不同的血细胞类型之间的定向分化,这些结果中的大部分结论可以通过其他文献中的结论进行佐证.分析表明,某些特定的单扰动或组合扰动可引起定向细胞分化.
此处介绍的整合网络和逻辑模型可以为HSC的分化选择提供良好的定性意见指导,也可以用于分析实现HSC转分化定性机制相关的影响因素.另外对于两个子网的整合过程也可以用作整合更多的造血分化和转分化调控规则的模板,以促进对造血体系的更全面的了解.此外,确定的扰动策略还可以为进一步的实验分析所需要确定的扰动目标提供一些意见.所有结论可以为血液病的研究与治疗提供建议.
猜你喜欢
上海师范大学学报·自然科学版(2022年3期)2022-07-11
农业工程学报(2022年6期)2022-06-27
现代电子技术(2022年11期)2022-06-14
清华金融评论(2022年4期)2022-04-13
汽车实用技术(2022年5期)2022-04-02
建材发展导向(2021年19期)2021-12-06
计算机仿真(2021年6期)2021-11-17
现代计算机(2021年10期)2021-05-28
湖南师范大学学报·自然科学版(2014年5期)2014-11-14
古代文明(2014年1期)2014-02-23
