
锌指蛋白调控机体炎症信号通路的研究进展
2022-04-15 07:17陈可添黎力之幸清凤廖晓鹏关玮琨文龙张海波郭冬生
中国畜牧杂志 2022年4期
陈可添,黎力之,幸清凤,廖晓鹏,关玮琨,文龙,张海波*,郭冬生*
(1.宜春学院生命科学与资源环境学院,江西省高等学校硒农业工程技术研究中心,宜春市功能农业与生态环境重点实验室,宜春学院继续教育学院,江西宜春 336000;2.江西正宇生物科技有限公司,江西宜春 336000)
1985 年,Miller 等首次发现转录因子IIIA 中锌指结构在基因调控方面起重要作用,并将含有锌指结构的蛋白命名为锌指蛋白。锌指蛋白基因敲除小鼠会逐渐出现恶病质、自身免疫性疾病、不完全性脱毛、关节炎和结膜炎等病症,锌指蛋白基因全身性敲除小鼠在出生后呈现生长迟缓甚至异常死亡、代谢功能障碍和严重低血糖等病况。外源注射10 mk/kg葡萄糖酸锌可显著增强大鼠体内锌指蛋白基因表达,降低骨髓中性粒细胞和二甲基亚砜诱导分化人原髓白血病细胞产生的肿瘤坏死因子-(TNF-),增强机体免疫力。Wu 等研究发现,急性肺损伤/急性呼吸窘迫综合征(ALI/ARDS)大鼠补充外源锌可上调锌指蛋白表达,显著降低TNF-、核因子-B(NFB)p65 的表达量,缓解脂多糖诱导的肺部炎症反应。NF-B 信号通路与Wnt/-catenin 信号通路在调控炎症反应过程中尤其关键,锌指蛋白对两者均有调节作用。其一,锌指蛋白通过扰乱NF-B 抑制蛋白激酶(IKK)和NF-B 抑制蛋白(IB)磷酸化,抑制受体相互作用蛋白1(RIP1)泛素化过程,阻碍NF-B 活化,影响其下游炎症通路;其二,锌指蛋白通过抑制糖原合酶激酶-3(GSK-3)磷酸化、与散乱蛋白(Dvl)蛋白竞争性结合支架蛋白(Axin),稳定-catenin 降解复合物完整性,调控炎症通路。本文就锌指蛋白介导动物机体炎症相关通路的调节作用进行综述,以期在调控动物炎症方面提供理论参考。
1 锌指蛋白简述
1.1 锌指蛋白概况 锌指蛋白是一类具有指状结构域的转录因子,用锌离子螯合剂除去锌指蛋白中锌离子或以Fe、Cu、Mn 等金属离子置换,均会降低锌指蛋白与DNA、RNA 等结合特异性,破坏蛋白结构稳定性。赵又佼等通过核磁共振发现锌指蛋白含锌离子时会折叠成紧密结构,锌离子夹叠在螺旋和两股反向平行的链之间;去除锌离子后发现锌指蛋白肽呈无规则卷曲,锌指序列无法正常形成,蛋白功能近乎完全丧失。锌指结构中有充分保守的区域,其结构稳定性主要由锌离子提供的链间交叉连接维持,其缺乏时“锌指”不能形成折叠结构,锌离子的存在使锌指蛋白呈现一个由不同肽序列折叠而成的完美螺旋构象。锌指蛋白因序列结构分不同构型,具有特异功能,如痘病毒锌指结构域(POZ)、CXXC 型锌指结构域(ZF-CXXC)等,锌指蛋白通过这些结构域特异性识别结合底物,从而调控靶基因表达,影响机体生理功能。
1.2 锌指蛋白分类 根据功能差异,锌指蛋白分为RING 蛋白、去泛素分子(DUBs)蛋白、CXXC 蛋白、POZ 蛋白等多个亚家族。RING 锌指蛋白具有进化上的保守性,与蛋白质互作调控体内靶基因表达。研究发现,转导通路相关蛋白和体内部分调节蛋白如乳腺癌敏感因子、转录中间因子TIF1、原癌蛋白CBL 等都含有RING 锌指区。DUBs 在泛素化途径中扮演重要角色,其主要通过水解泛素羧基末端酯键、肽键或异肽键,将泛素分子特异性从链接泛素蛋白或前体蛋白中释放。CXXC 锌指蛋白家族特有基序ZF-CXXC,由两对保守富含胞嘧啶集团结合2个锌离子组成,可特异性识别并结合富含CpG 二核苷酸的DNA 序列。研究表明,人类基因5q31 染色体区缺失型锌指蛋白基因易发生急性髓性白血病和骨髓增生异常综合征等疾病,该家族内如锌指蛋白CXXC4、锌指蛋白CXXC5除信号传导、细胞凋亡、细胞能量代谢和血管生成等生物学功能外,还介导Wnt/-catenin 信号通路调控机体免疫。锌指蛋白ZBTB 亚家族属于POZ 蛋白家族,该亚家族成员N 端均有一个BTB/POZ 结构域,用以调节蛋白间相互作用;C 端数个CH锌指结构域,介导蛋白与DNA 序列间的识别、结合作用。该亚家族成员具有转录抑制功能,如锌指蛋白ZBTB20 抑制产生Toll 样受体4(TLR4)触发的巨噬细胞前炎性细胞因子和干扰素(IFN-),调节机体免疫功能。总之,RING 蛋白、DUBs 蛋白、CXXC 蛋白、POZ 蛋白等锌指蛋白亚家族本同末异,在NF-B、Wnt/-catenin 通路等途径中调节机体免疫。
2 锌指蛋白对动物炎症通路的影响
2.1 锌指蛋白对NF-B 通路的调节作用 正常生理条件下,NF-B 以一种无生理活性的二聚体形式存在于胞浆。当机体受到炎性细胞因子或其他外来刺激时,诱导胞质内IB 磷酸化、泛素化而降解,导致胞内NF-B增多,诱使其下游炎性细胞因子合成释放,机体发生炎症反应、细胞凋亡等组织损伤现象。锌指蛋白A20是炎性反应的内源性调控蛋白和组织细胞保护性蛋白,在B 细胞、T 细胞和成纤维细胞中均表达,对NF-B负性调控功能体现在减轻机体炎症反应及缓解免疫损伤方面。锌指蛋白A20 作为DUBs 家族中的一员,其N 端含有半胱氨酸蛋白酶/DUB 结构域OUT,C 端含有7个锌指结构,其中第4 锌指有E3 泛素连接酶活性,具有去泛素化和泛素化活性的双重功能。一方面,锌指蛋白A20 通过N-末端脱泛素因子干扰肿瘤坏死因子受体相关因子6(TRAF6)、RIP1、NF-B 必需调制蛋白(NEMO)等泛素化修饰,抑制下游IB 磷酸化;另一方面,锌指域C-末端E3 泛素连接酶泛素化修饰TLR、核苷酸结合寡聚化结构域蛋白2(NOD2)、T 细胞抗原受体(TCR)和B 细胞抗原受体(BCR),抑制NF-B 信号通路。
随着研究的深入,现已确定凡能激活NF-B 信号通路的因素均能诱导锌指蛋白A20 表达上调。研究发现10 ng/mL TNF-处理大鼠髓核细胞72 h 后,锌指蛋白A20、p53、p-p65 等表达均显著提升。急性肝损伤模型小鼠血清和肝组织中TNF-表达较正常组小鼠升高,肝组织白细胞介素1()、mRNA表达及p-IB和p-NF-B p65 蛋白表达均增加;外源药物增强锌指蛋白基因表达后发现小鼠血清和肝组织TNF-浓度降低,肝组织NF-B p65 蛋白升高,mRNA 及p-IB和p-NF-B p65蛋白表达显著降低。TNF-与受体TNFR2 结合后使其汇集,泛素化RIP1 的K63 位,激活转化生长因子激酶并与IKK-结合形成复合体,IB发生泛素化,引起NF-B 进入细胞核,激活下游靶基因。上调锌指蛋白表达可抑制NF-B 信号通路,调控炎症反应。试验发现,在TNF 激活的NF-B 信号通路中,锌指蛋白A20、多聚泛素化链和NEMO 形成复合物,利用锌指蛋白A20 去泛素化功能限制IKK 活化,以及锌指结构域7(ZnF7)与K63 多聚泛素化链结合共同作用使RIP1 被蛋白酶降解;同时,锌指蛋白A20 还可使TRAF2 泛素化降解,抑制NF-B 信号活化。锌指蛋白A20 还可阻碍NF-B 作用于IL-1、CD40 的反应,抑制模式识别受体发出的信号及T 细胞、B 细胞抗原受体活化。锌指蛋白ZBTB20,又称ZNF288、DPZF或HOF,是BTB/POZ 家族的独特成员,主要通过N端BTB/POZ 结构域和C 端CH保守的Krüppel 型锌指作为转录抑制因子发挥作用,在机体脾脏、胸腺、淋巴结等组织器官广泛表达。Liu 等特异性敲除小鼠髓系细胞锌指蛋白基因,发现多种TLR 配体反应性下降,巨噬细胞产生的炎性细胞因子如TNF、IFN-、IL-6 等减少。另外,锌指蛋白ZBTB20 通过抑制TLR 下游信号通路中基因转录,下调NF-B表达。综上所述,锌指蛋白对机体NF-B 的调控机理主要通过下调基因表达,干扰IB 磷酸化,抑制NF-B 活化,减少促炎因子分泌,平衡机体炎症反应(图1)。
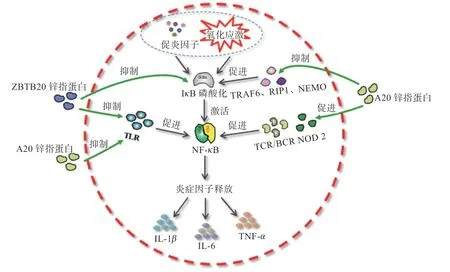
图1 锌指蛋白介导NF-кB 信号转导通路调节动物肠道炎症
2.2 锌指蛋白对Wnt/-catenin 信号通路的影响 Wnt/-catenin 信号通路具有进化保守、高度复杂的特点,由配体蛋白Wnt 和膜蛋白受体结合激发信号转导通路,调节器官发育、组织稳态及损伤修复等重要过程。外界刺激(如损伤、病原体、激动剂等)使Wnt 信号通路异常,抑制GSK-3活性,导致细胞内-catenin 聚集过多,与T 细胞因子/淋巴增强因子(TCF/LEF)相互作用,引起机体过度炎症反应。如发生骨关节炎时,Wnt/-catenin 信号通路过表达抑制软骨细胞增殖及II型胶原蛋白分泌导致软骨退化,另外还使滑膜成纤维细胞过度增殖,及I 型胶原沉积引起滑膜炎症。多发性硬化(中枢神经系统炎性脱髓鞘性病变)大鼠模型体内Wnt3、p-GSK-3、-catenin 蛋白水平显著升高,模型鼠脊髓呈现大量炎性细胞浸润,TNF-、IL-1等炎症因子浓度升高。
锌指蛋白CXXC 家族调控Wnt/-catenin 信号通路,一是当机体处于稳定状态时,通过降低GSK-3磷酸化,使-catenin 降解保持低值;同时,锌指蛋白CXXC 过表达抑制Wnt/-catenin 信号通路关键蛋白,竞争性拮抗使-catenin 降解复合物免受破坏。在Wnt 配体未激活信号通路时,Axin、腺瘤样息肉蛋白(APC)、GSK-3和酪蛋白激酶(CK I)组成复合体,使-catenin 磷酸化后被-转导素重复序列包含蛋白泛素化降解。对胶原蛋白II 诱导的类风湿关节炎大鼠注射GSK-3抑制剂后发现,前列腺素E2,5-羟色胺和组胺等自身活性物质增加,血清中IL-6、IL-12 和TNF-等细胞炎性因子显著增多。Giacoppo 等建立实验性变态反应性脑脊髓炎小鼠模型发现,GSK3-活性受到抑制,-catenin上调,入核后调节下游基因表达。正常状态下锌指蛋白CXXC4 对GSK-3有保护作用,可抑制GSK-3的磷酸化。李萍等研究发现,锌指蛋白基因甲基化异常的细胞组中p-GSK-3、Wnt、-catenin 相对量均显著高于正常组,表明当锌指蛋白CXXC4 基因被抑制时,GSK-3磷酸化加剧,Wnt/-catenin 通路上调。
当机体发生炎症反应或受到外界刺激时,Wnt 配体蛋白与卷曲蛋白(FZD)、低密度脂蛋白受体相关蛋白5/6(LRP5/6)结合,形成Wnt-FZD-LRP5/6 三聚体复合物,使细胞内Dvl 蛋白活化并向质膜迁移。研究表明,锌指蛋白CXXC4、CXXC5 能作为Dvl 蛋白结合伴侣,负反馈调节Wnt/-catenin 信号通路。锌指蛋白CXXC4 通过其特殊的“KTXXXI”基序直接与Dvl 蛋白PDZ 结构域相互作用,稳定-catenin 降解复合物。锌指蛋白CXXC5 与Dvl 蛋白相互作用在多种细胞内被发现,如成骨细胞、皮肤细胞、神经干细胞等。Lee 等在皮肤损伤后修复愈合的研究中发现,锌指蛋白CXXC5 含有C-末端的Dvl 结合结构域,其与Dvl 相互作用,成为Wnt/-catenin 途径的负反馈调节因子。对生长期小鼠肺成纤维细胞分别添加目的基因腺病毒载体、基因抑制载体感染48 h后发现,添加组-catenin、Dvl 蛋白量显著减少,抑制组-catenin、Dvl 蛋白表达量显著增加;此外,在TGF-1 刺激下细胞-catenin、Dvl 蛋白表达增加,添加基因载体后,两者逐渐减少。表明锌指蛋白CXXC5 在活化的Wnt/-catenin 信号通路中,过表达影响Dvl 蛋白含量调控-catenin 浓度。综上所述,当Wnt/-catenin 信号通路无Wnt 配体存在时,锌指蛋白通过抑制GSK-3磷酸化,稳定-catenin 降解复合物,抑制下游IL-1、TNF-等炎症因子分泌;当Wnt 配体激活Wnt/-catenin 信号通路,锌指蛋白与Dvl 蛋白进行竞争性结合,阻碍Dvl 蛋白与Axin-GSK-3作用,间接稳定-catenin 降解复合物,阻断炎症过度进程(图2)。
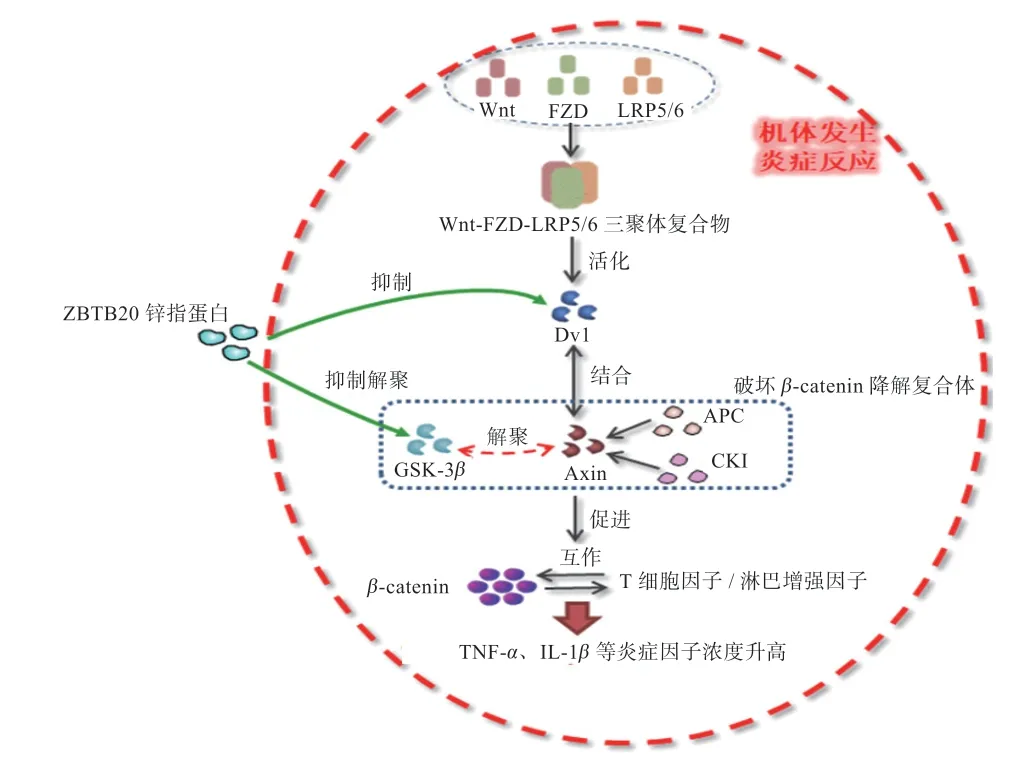
图2 锌指蛋白介导Wnt/β-catenin 信号转导通路调节动物炎症
3 总结与展望
在基因炎症通路中,锌指蛋白可以调控基因表达,干扰IB磷酸化,抑制NF-B 信号通路,减少炎症因子分泌。在Wnt/-catenin 信号通路中,锌指蛋白会抑制GSK-3磷酸化,保持-catenin 降解复合物完整性;另外,锌指蛋白还会与Dvl 蛋白竞争,保护Axin 蛋白,调控Wnt/-catenin 通路,抑制炎症反应。目前对锌指类蛋白的研究主要集中在分析锌指蛋白结构,建立其与DNA 互作模型。但是关于锌指蛋白的研究仍存在不足。第一,锌指蛋白家族庞大种类较多,除对C2H2 型研究较透彻外,其他各类型的结构功能有待深入探索。第二,锌指蛋白功能研究大多集中在机体免疫炎症抗肿瘤等方面,在应对一些非生物胁迫如热应激等环境下相关研究较少。随着对锌指蛋白结构和功能研究的不断深入,阐明锌指蛋白调控机理,人工锌指蛋白技术将具有更广阔的应用前景。
猜你喜欢
中国现代医生(2022年19期)2022-11-04
中国现代医生(2022年19期)2022-08-25
中国典型病例大全(2022年11期)2022-05-13
祝您健康(2018年10期)2018-10-11
中文信息(2017年2期)2017-04-13
江苏农业科学(2016年11期)2017-03-21
智能计算机与应用(2016年4期)2016-09-26
天津农业科学(2009年1期)2009-12-28
销售与管理(2006年9期)2006-09-17
