
高原鼠兔干扰对川西北高寒草甸植物群落及土壤物理性状的影响
2022-05-11 10:44根呷羊批周俗杨孔王钰杨思维刘刚杨廷勇扎德王泽光
草原与草坪 2022年1期
根呷羊批,周俗,杨孔,王钰,杨思维,刘刚,杨廷勇,扎德,王泽光
(1.西南民族大学青藏高原研究院,四川 成都 610041;2.四川省甘孜藏族自治州动物疫病预防控制中心,四川 康定 626000;3.四川省林业科学研究院,四川 成都 611731;4.四川省草原科学研究院,四川成都 610041;5.甘孜藏族自治州草原工作站,四川 康定 626000;6.若尔盖县科学技术和农业畜牧局,四川 若尔盖 624500;7.若尔盖县林业和草原局,四川 若尔盖 624500)
草原是畜牧业可持续发展的生态和物质保障,近年来,高寒草甸的健康状况开始恶化,造成严重的社会和生态问题[1]。其中,鼠害问题尤为突出,2019年四川省林业和草原局发布的四川省草原监测报告显示,2018年全省草原鼠害发生面积约990.93万hm2,草原生物灾害发生严重,高原鼠兔等鼠类的活动与高寒草甸草地生态环境问题日益突出,直接威胁着青藏高原草地畜牧业的可持续发展[2],主要表现在植株高度降低、植物种类减少,产草量低下、土壤肥力下降等。因此,研究川西北高寒草甸退化机理,对于治理和恢复退化高寒草甸具有重要意义[3]。
高原鼠兔干扰对高寒草甸具有正面和负面的双重影响[4],正面影响是高原鼠兔作为高寒草甸关键物种,成为了冬季草原上许多肉食动物的唯一食物来源。此外,其挖掘的洞穴可以为许多小型鸟类和蛙类提供赖以生存的巢穴,挖掘行为亦可达到疏松土壤的作用,通过加速土壤循环过程对生态系统产生正效应,对微生境造成干扰,从而引起植物多样性的增加,对地表及地下(根部)生物量也有积极意义[5]。负面影响是高原鼠兔挖掘活动导致微地形结构变化,表现为土壤肥力下降[6],植物群落内不可食牧草急剧增加[7],原有优势植物高山嵩草(Kobresiapygmaea) 种群的生态位明显衰减[8],甚至引起水土流失加剧,扩大裸地面积,破坏草地结构[10],威胁高寒草甸生态系统的健康。
有关高原鼠兔对高寒草甸的影响的报道较多,但结果不尽相同。孙飞达等[10]在青海省果洛州的研究发现随高原鼠兔密度增加,高寒草甸植物物种群落多样性指数、均匀度指数呈现先降低后增加的趋势,当高原鼠兔在中等密度时,群落多样性的指标值均为最小。Pang等[11]也发现物种均匀度指数随高原鼠兔干扰程度增加而呈先增加后降低趋势,而贾婷婷[12]在甘南玛曲县的研究发现,随高原鼠兔干扰水平的增加,植物群落盖度、丰富度指数、多样性指数先升高后降低,在中等干扰强度下群落多样性各指标值均为最大;而王莹等[1]在甘南藏族自治州玛曲县的研究却有不同的结果,随高原鼠兔干扰程度增加,高寒草甸植物丰富度指数、均匀度指数以及多样性指数呈降低趋势,土壤全氮含量随高原鼠兔干扰水平增加而降低,张卫红等[14]在西藏邦杰塘高寒草甸也得出了类似的研究结果。而严红宇等[15]发现,相同高原鼠兔密度的栖息地内杂草斑和秃斑植物总盖度、平均高度和总地上生物量没有显著差异。此外,田富等[16]研究表明高原鼠兔干扰对高寒草甸植被群落多样性指数影响并不强,不同干扰强度下的高寒草甸植物多样性指数没有显著差异。
综上所述,关于高原鼠兔对高寒草地的影响研究主要集中在青藏高原地区[17],其中以青海省三江源自然保护区[6-10]和甘肃省甘南草原[7,18]为主。然而,川西北高原作为青藏高原重要组成部分,也是中国5大牧区之一,具有重要的生态价值。针对高原鼠兔对川西北高寒草甸植物群落特征及土壤影响的相关研究甚少,且植被群落特征和土壤理化性状对高原鼠兔扰动的响应研究较少。因此,本研究以川西北高原若尔盖高寒草甸为例,通过分析高原鼠兔干扰及不同干扰强度下高寒草甸的植被盖度、高度、多样性指数,以及土壤物理性状等,探讨高原鼠兔对高寒草甸的扰动作用,以期为正确认识高原鼠兔在川西北草地生态系统中的作用和川西北草原鼠荒地恢复治理提供科学依据。
1 材料和方法
1.1 研究区概况
研究区位于青藏高原东部边缘的若尔盖境内(E 102°08′~103°39′,N 32°56′~34°19′)(图1),植被类型以高寒草甸为主,海拔3 476 m,该地区气候为典型高原寒带湿润季风气候,冬长无夏,年平均气温0.7 ℃,年均降水量663.6 mm。土壤类型为高山草甸土,其建群种为矮嵩草(Kobresiahumilis),主要伴生种有垂穗披碱草(Elymusnutans)、鹅绒委陵菜(Potentillaanserina)、黄帚橐吾(Ligulariavirgaurea)等。试验区为冬季草场,放牧压力较轻,小型哺乳类动物中高原鼠兔为绝对优势种,偶见高原鼢鼠(Eospalaxfontanierii)土丘。
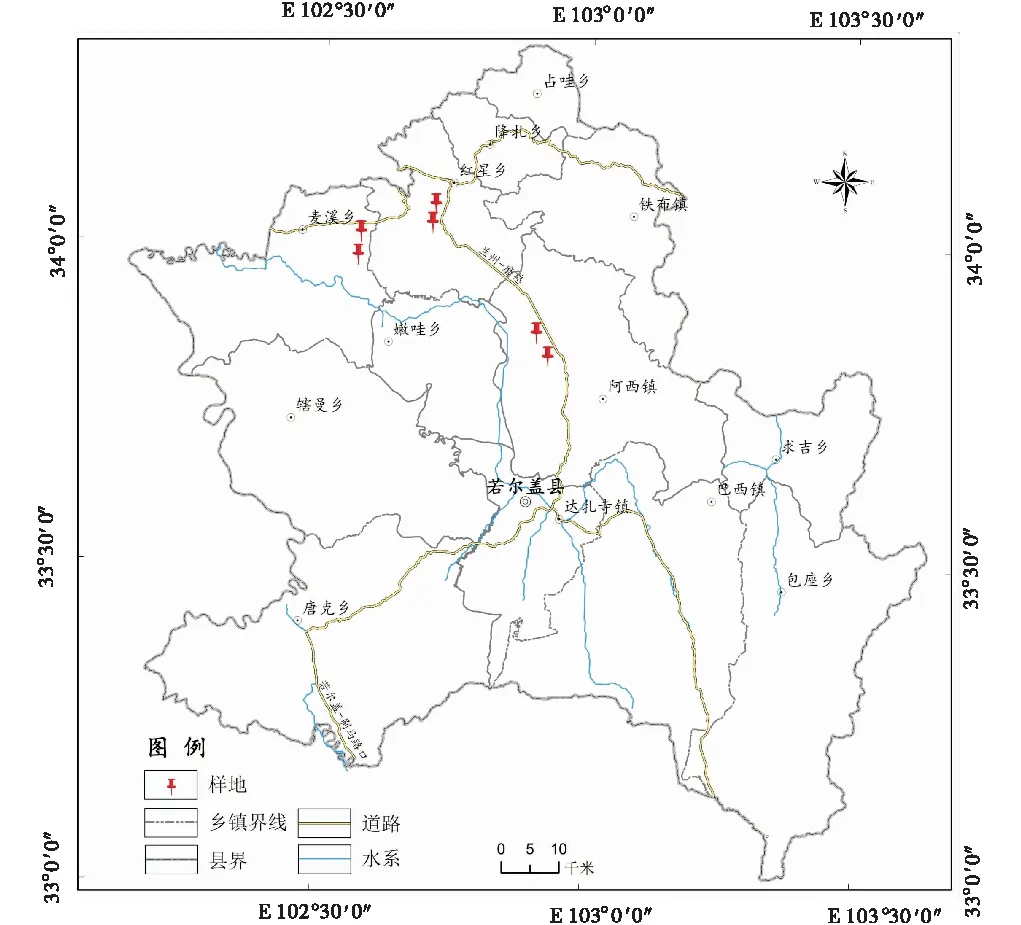
图1 研究区位置示意图Fig.1 Location of the study area
1.2 试验方法
1.2.1 试验设计 于2019年7月盛草期进行野外调查,为减少放牧因素的影响,选择地势平坦一致且放牧压力较轻的冬季草地设置样地。高原鼠兔干扰区与非干扰区有时是渐进的,有时是镶嵌分布的。本研究高原鼠兔干扰区选择以现场观察是否有高原鼠兔洞口和高原鼠兔出没为准,若同时发现有高原鼠兔洞口和高原鼠兔出没,则选定为高原鼠兔干扰区。在高原鼠兔干扰区共设置15个大小为1/4 hm2的鼠兔密度调查圆形(半径为28.2 m)样地,连续3 d用“堵洞盗洞法”进行高原鼠兔密度调查[19-20],采用有效鼠洞口数来指示高原鼠兔干扰强度[21-22],15个干扰样地按有效洞口数从低到高依次用B1-B15表示(表1)。然后,在干扰区附近选择无高原鼠兔侵扰的同等大小样地10个,且干扰区和非干扰区的距离大于500 m。同时,在干扰区和非干扰区调查各样地植被及土壤指标,每个样地重复3次。

表1 干扰区样地有效洞口数
1.2.2 植被样方调查 以每块样地为中心设置1个样方(1 m×1 m),向外任意两方向延伸一定距离再设置2个样方,每个样地共计3个样方,呈V字状,记录每个样方内的物种数,测定样方内所有植物的高度(每种植物随机测20株自然高度,取平均数)、总盖度(目测法)、分盖度(针刺法测定)、多度(记录每个物种的个数作为物种多度)和频度(样地内随机扔频度圈30次,记录物种出现频率)。根据青藏高原已有植物功能群的划分实例[23],本研究依据不同植物的生活型,将其分别划分为禾草科(Graminea)、莎草科(Cyperaceae)、豆科(Leguminosae)和杂类草4个功能群。
1.2.3 土壤调查 进行植被调查的同时,在每个样方附近选取3个不同的采样点,且避开鼠兔裸露斑块,样点间距大于5 m,用直径为5 cm的土钻采集表层(0~20 cm)土壤样品,用紧实度仪(SC500)测定表层(0~20 cm)和深层(20~40 cm)土壤紧实度。
1.3 数据处理与分析
1.3.1 土壤容重和含水量 土壤容重(di)采用环刀法测算[24],计算公式为:
土壤含水量采用烘干法测算。计算公式为:
式中:W为含水率(%);W1为环刀重(g);W2为称环刀+湿土重(g);W3为环刀+烘干土重(g)。
1.3.2 重要值 重要值是计算物种多样性指数的重要指标,本研究采用相对分盖度、相对高度和相对频度计算物种相对重要值[25-26]。
Pi=[(Ci/C+Fi/F+Hi/H)]×100%
式中:Pi为1 m2样方中第i种的重要值;Ci表示i种在样方中的分盖度,Fi表示i种在单个样方中出现的次数,Hi表示i种的平均高度;C、F、H分别表示样方中物种的总盖度、出现的总次数、总平均高度。
1.3.3 物种多样性 采用物种丰富度指数、多样性指数、均匀度指数和辛普森指数表示物种多样性[27-29]。其中,物种丰富度指数(S)采用Patrick指数;多样性指数采用Shannon-Wiener指数(H)和辛普森指数(D),均匀度指数(J)采用 Pielou,植物功能群多样性的测度采用Shannon-Wiener指数(H′),具体计算公式如下:
S=样方内出现的物种总数
J=H/Ln(S)
式中:Pi是种的相对重要值,H′为功能群多样性指数;S′为样方内该功能群中所包含的物种总数,Pi′是指该功能群内所有种的相对重要值。
1.4 统计分析
采用Excel 2016.0进行数据的基本统计、采用独立样本T检验分析高原鼠兔干扰对测定指标的影响,若差异不显著,则不做进一步分析,若差异显著(P<0.05),则对不同高原鼠兔密度下各指标的变化进行One-way ANOVA分析,两两比较采用LSD检验,选用各组平均数±标准误,数据分析用SPSS 21.0统计软件,并借助Origin 2018.0绘图。
2 结果与分析
2.1 高原鼠兔干扰对高寒草甸植被平均高度和总盖度的影响
高原鼠兔干扰对植被平均高度、总盖度的影响一致,表现为高原鼠兔干扰显著降低了地上植被盖度和总盖度(F=56.391,P<0.05,n=75;F=13.422,P<0.05,n=75)(图2)。
随着高原鼠兔密度增加,植被平均高度在样地B1-B15,从最高为B1(135个/hm2)的17.7 cm降到最低为B11(960个/hm2)的15.6 cm,植被总盖度从B3(135个/hm2)的最大值78.4%下降至B15(1 215个/hm2)的最低值31.5%。总体上看,地上植被平均高度和总盖度均随干扰强度增强呈下降的趋势(图3)。
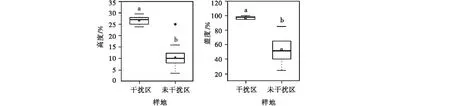
图2 高寒草甸植被平均高度、总盖度Fig.2 Effect of plateau pika disturbance on vegetation height and coverage in alpine meadow

图3 高原鼠兔干扰区不同干扰强度下的植被高度、盖度Fig.3 Effect of plateau pika disturbance on vegetation height and coverage in alpine meadow注:不同小写字母表示差异显著(P<0.05)。下同
2.2 高原鼠兔干扰对高寒草甸植物物种多样性的影响
高原鼠兔干扰对植物物种丰富度指数和均匀度指数的影响一致,具体表现为高原鼠兔干扰显著降低了物种丰富度指数(F=0.006,P<0.05,n=75)、均匀度指数(F=0.200,P<0.05,n=75),但显著增加了Shannon指数(F=5.279,P<0.05,n=75),对辛普森指数无显著影响(F=0.183,P=0.079>0.05,n=75)(图4)。

图4 高原鼠兔干扰下的高寒草甸物种多样性Fig.4 Effect of plateau pika disturbance on species diversity in alpine meadow
随着高原鼠兔干扰强度增加,丰富度指数在B7(465个/hm2)达到最高值,为24种,在B14(1 080个/hm2)最低,为8种;Shannon指数在B8(495个/hm2)达到最高值2.41,而在B15(1 215个/hm2)降至最低值1.17,均匀度指数在B8(495个/hm2)达到最高值0.84,而在B14(1 080个/hm2)降至最低值0.46。总体上丰富度指数、Shannon指数和均匀度指数变化趋势相一致,呈先增加后降低的趋势(图5)。
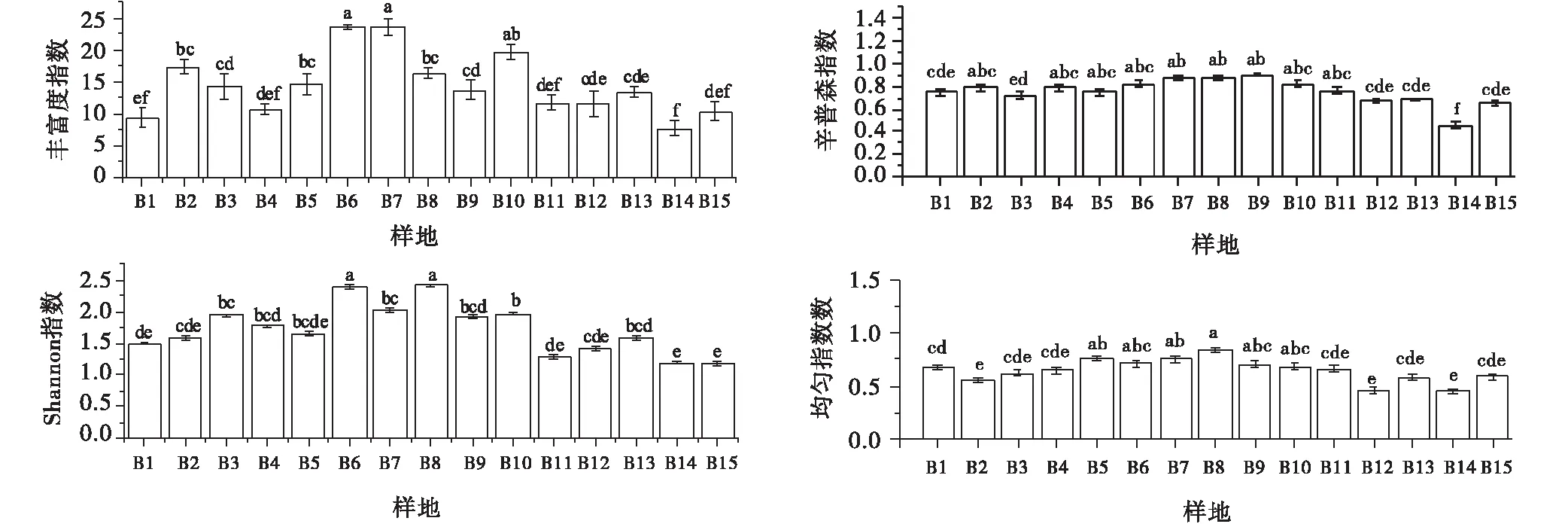
图5 高原鼠兔干扰区不同干扰强度下高寒草甸物种多样性Fig.5 The species diversity under different disturbance intensities of plateau pika
2.3 高原鼠兔干扰对高寒草甸植物功能群多样性的影响
除莎草科功能群多样性指数(F=10.355,P=0.579>0.05,n=75)外,高原鼠兔的干扰显著降低了禾本科功能群多样指数(F=13.129,P<0.05,n=75)、豆科功能群多样性指数(F=12.228,P=0.004<0.05,n=75),而显著增加了杂类草功能群多样性指数(F=0.204,P<0.05,n=75)(图6)。

图6 高原鼠兔干扰下的高寒草甸植物功能群多样性Fig.6 Effect of plateau pika disturbance on functional diversity in alpine meadow
随着高原鼠兔干扰强度增加,禾本科、莎草科和杂类草功能群多样性指数呈不规则的趋势,豆科功能群多样性指数呈下降的趋势。其中,禾本科多样性指数B3(270个/hm2)最高,为0.48,B13(1 050个/hm2)达最低,为0;莎草科多样性指数B9(735个/hm2)达最高,为0.69,B5(330个/hm2)最低,为0.19;豆科多样性指数B1(135个/hm2)最高,为0.53,B15(1 215个/hm2)最低,为0;杂类草多样性指数B9(735个/hm2)最高,为1.60,B5(285个/hm2)最低,为0.55(图7)。

图7 高原鼠兔干扰区不同干扰强度下高寒草甸植物功能群多样性Fig.7 The functional diversity under different disturbance intensities of plateau pika
2.4 高原鼠兔干扰对高寒草甸植物群落物种组成及其重要值的影响
高原鼠兔的干扰显著降低了禾本科功能群重要值(F=9.855 4,P<0.05,n=75),其中,垂穗披碱草、发草的重要值显著下降,而增加了异针茅的重要值;高原鼠兔的干扰显著降低了莎草科功能群重要值(F=9.855,P<0.05,n=75)和豆科功能群重要值(F=90.117,P<0.05,n=75),其中,矮嵩草重要值有所上升,而青藏薹草重要值显著下降,此外,黄花棘豆和多枝黄耆重要值显著下降;高原鼠兔的干扰显著增加了杂类草功能群重要值(F=4.140,P<0.05,n=75),其中鹅绒委陵菜、冷蒿、车前草等植物的重要值显著上升(图8,表2)。

图8 高原鼠兔干扰下的高寒草甸植物功能群重要值Fig.8 Effect of plateau pika disturbance on functional important value in alpine meadow

表2 高原鼠兔干扰下高寒草甸植物群落的物种组成及其重要值
2.5 高原鼠兔干扰对高寒草甸土壤紧实度和容重的影响
高原鼠兔的干扰显著影响了高寒草甸土壤紧实度、容重和含水量,具体表现为高原鼠兔干扰下显著降低了土壤紧实度(0~20 cm土层:F=0.028,P<0.05;22.5~40 cm层:F=0.028,P<0.05)和土壤含水量(F=45.635,P<0.05),但显著增加了高寒草甸的土壤容重(F=56.579,P<0.05)(图9)。
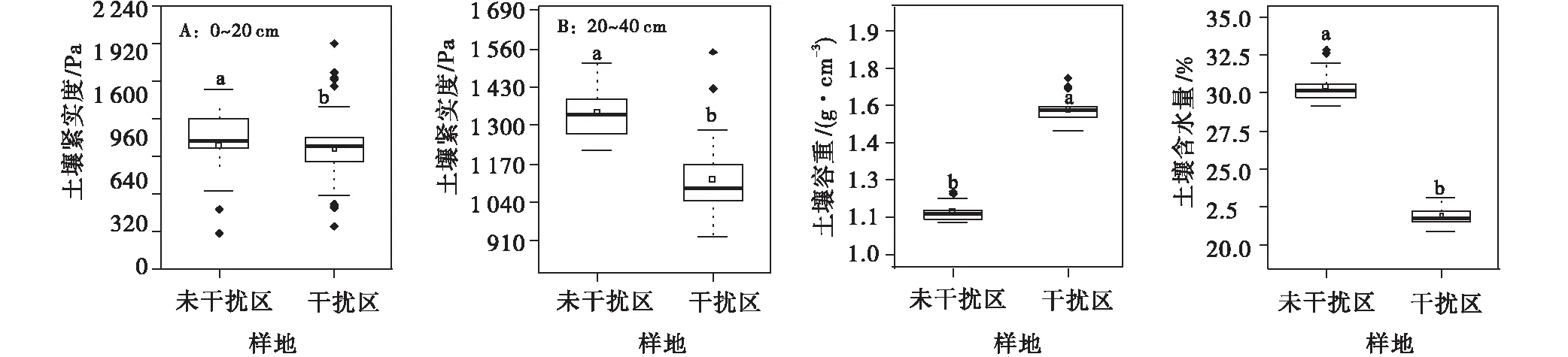
图9 高原鼠兔干扰下的土壤物理性状Fig.9 Effect of plateau pika disturbance on soil physical properties in alpine meadow

图10 高原鼠兔干扰区不同干扰强度下高寒草甸土壤物理性状Fig.10 The soil physical properties under different disturbance intensities of plateau pika
随着高原鼠兔干扰强度加重,表层土壤紧实度、深层土壤紧实度、土壤容重和含水量变化明显。其中,0~20 cm土层土壤紧实度B8(495个/hm2)达到最大,为1 522.88 Pa,20~40 cm层土壤紧实度B2(165个/hm2)最大,为1 314.26 Pa;土壤容重B13(1 050个/hm2)达到最大,为1.69 g/cm3,而土壤含水量呈先上升后下降趋势,B9(735个/hm2)最大,为32.75%。
3 讨论
3.1 高原鼠兔干扰对高寒草甸植物群落的影响
高原鼠兔是青藏高原的重要物种,且种群数量巨大[30],其采食行为不仅会直接影响高寒草甸植物生长发育,还通过挖掘土壤、排泄粪尿等活动间接影响植物生长发育[31-32]。本研究结果显示,高原鼠兔干扰区植被高度显著低于未干扰区植被高度,且随着干扰强度增加样地内植被高度总体呈下降趋势,据刘伟等[33]研究发现植物平均高度的降低与高原鼠兔理想栖息地环境有关,它们将较高植物视为一种风险源,因而对较高植物进行刈割,降低植物平均高度。而据宋梓涵等[34]报道,高原鼠兔对栖息地植被条件选择具有一定的倾向性,其主要选择植被高度较低、视野开阔、地势平坦的草地;高原鼠兔侵扰的草地植被高度呈现出整体较低的原因既有可能是高原鼠兔活动造成的[35],也有可能是高原鼠兔对栖息地选择的结果[36]。此外,高原鼠兔干扰显著降低了植被盖度,且随着干扰强度增加植被盖度呈下降趋势,表明高原鼠兔啃食植物和挖掘土壤行为造成草地出现破碎化的秃斑地或次生裸地的景观,群落处于次生演替阶段,草地鼠丘的形成也减少了高寒草甸可利用面积,这与刘季科等[37]对高原鼠兔数量与危害程度的关系研究相一致,与刘伟等[38]在青海省果洛研究高原鼠兔对退化草地植物群落结构及恢复演替的影响结果相一致。
高原鼠兔干扰显著降低了物种丰富度指数和均匀度指数,显著增加了多样性指数,随干扰强度增加,物种丰富度指数、均匀度指数和多样性指数均呈先上升后下降趋势,且在高原鼠兔洞口密度375~735个/hm2段达到最高,刘菊梅等[9]也认为适当的鼠兔种群密度对保持草地植被生物多样性有一定的积极作用。从植被不同功能群分析,高原鼠兔干扰显著降低了禾本科和豆科功能群多样性指数,而显著增加了杂类草功能群多样性指数,对莎草科功能群多样性指数没有显著影响,随干扰强度增加,禾本科和莎草科功能群多样性指数没有显著变化规律,豆科功能群多样性指数呈下降趋势,杂类草功能群多样性呈先上升后下降变化趋势,主要原因是高原鼠兔干扰增加了高寒草甸生境的异质性,为不同物种共存提供了条件,从而增加了植物多样性指数[39]。对植被功能群进一步研究发现,高原鼠兔干扰也显著降低了莎草科、禾本科和豆科重要值,而显著增加了杂类草重要值。通过对比研究干扰区与未干扰区不同种群植物重要值发现,矮嵩草重要值有所上升,而青藏薹草重要值显著下降,此外,黄花棘豆和多枝黄耆重要值显著下降,而鹅绒委陵菜、冷蒿、车前草的重要值显著上升,造成这一结果的原因可能与高原鼠兔的食性有关。据康宇坤等[40]研究表明,高原鼠兔在不同时期的食性及采食同种植物的比例都有所差异,且在盛草期主要采食对象为禾本科(垂穗披碱草)、莎草科(矮嵩草)、蔷薇科(鹅绒委陵菜),也有研究显示棘豆属植物是高原鼠兔的重要食物,占其天然饲料的很大比例[41],这一结果与金少红[42]、田富等[43]的研究结果一致。
3.2 高原鼠兔干扰对土壤物理性质的影响
土壤作为植被生长所需养分与水分的来源,是植被生长发育的物质基础,为作物提供必需的生活条件[44]。其中,土壤含水量对高寒草甸气候变化产生着重要影响,而影响高寒草甸土壤含水量的主要因素有季节性降水、太阳照射量,地表温度、土壤蒸发、植物蒸腾、以及家畜放牧和啮齿类动物(高原鼠兔等)干扰等。川西北高寒草甸每年降水量基本保持在600 mL左右,且主要6-8月,土壤含水量成为了物种多样性,动植物生长的重要环境影响因子之一[45],对维持生物多样性也起着重要的支撑作用[46],同时土壤含水量对啮齿动物干扰后高寒草甸植被生长和恢复也起着关键作用[47]。此外,与土壤含水量对应的土壤容重和土壤紧实度,也是表征土壤物理性状的重要指标。本研究通过研究高原鼠兔干扰对川西北高寒草甸土壤物理性质的影响,发现高原鼠兔干扰显著降低了高寒草甸土壤含水量和紧实度,但显著增加了表层土壤容重。随干扰强度的增加,表层土壤的含水量呈明显的先上升后下降趋势,与之相对应的土壤容重随干扰强度增加呈先下降后上升的趋势,土壤紧实度没有显著变化规律。表明高原鼠兔通过啃食地面植物,使地表裸露面积增大,以及通过挖掘土壤将养分较高的表层土壤翻抛至地表,经风蚀、水蚀等作用,使得土壤肥力及水分大量损失[48],导致表层土壤容重增加,引起土壤持水能力下降,且表现为干扰强度越大,土壤水分含量越少,与朱灿等[49]在若尔盖和孙飞达等[50]在三江源区相关研究结果一致。
4 结论
1)高原鼠兔干扰显著降低了地上植被盖度、高度、植物物种丰富度指数、均匀度指数、土壤紧实度和土壤含水量、禾本科和豆科功能群多样性和重要值,但显著增加了土壤容重和杂类草功能群多样性和重要值。
2)随干扰强度增强,地上植被平均高度、总盖度、豆科功能群多样性指数均呈下降的趋势;随干扰强度增强,丰富度指数、Shannon指数、均匀度指数和土壤含水量呈先增加后降低的变化趋势。
3)适度的鼠兔干扰,增加了高寒草甸物种多样性和土壤含水量,高原鼠兔有效洞口密度为375~735个/hm2是其在高寒草甸的最适密度。
猜你喜欢
中国交通信息化(2022年7期)2022-10-27
课外生活·趣知识(2021年2期)2021-05-24
海外文摘·文学版(2021年3期)2021-05-19
学校教育研究(2021年24期)2021-03-28
西部资源(2018年3期)2018-11-06
湖北畜牧兽医(2017年4期)2017-05-18
绿色科技(2017年7期)2017-05-12
延河·绿色文学(2016年8期)2016-05-14
纺织导报(2014年8期)2014-10-31
