
色季拉山不同环境梯度冷杉群落木本植物分布格局及功能多样性研究
2022-05-25 03:24汪汉驹李江荣郑维列陈文盛丁慧慧
绿色科技 2022年8期
汪汉驹,李江荣,郑维列,陈 康,陈文盛,丁慧慧
(1.西藏农牧学院高原生态研究所,西藏 林芝 860000;2.西藏高原森林生态教育部重点实验室,西藏 林芝 860000;3.西藏林芝高山森林生态系统国家野外科学观测研究站,西藏 林芝 860000)
1 引言
了解沿环境梯度的植被格局变化是生态学中一个长期存在的问题,预测环境因素如何推动群落构建仍然是一个重大挑战[1]。量化物种功能多样性是完成这项工作的关键之一。
植物功能性状是指能够强烈影响生态系统功能以及能够反映植被对环境变化响应的核心植物属性,这些属性是大多数植物具有的共有或常见性状[2,3]。功能性状对植物体定植、存活、生长和死亡存在潜在一系列显著影响,同时这些属性能够单独或联合指示生态系统对环境变化的响应,并且能够对生态系统过程产生强烈影响[4]。
目前,基于功能性状的研究方法越来越多地应用于群落生态学当中[5]。物种的功能性状中,植株最大高度(Maximum height,HMAX)不仅影响植物对光照、水分和热量的利用,而且影响植物种子的扩散,它代表了光获取和树干生长成本之间的权衡。叶面积(Leaf area,LA)与水分利用率有关,因此LA低的个体通常具有更好的耐寒耐旱性。LA还与光合能力有关,LA高的物种在阴暗条件下能获取更多的光。高LA既是一种避荫(增加生长速率)策略,也是一种耐荫(优化光的捕获)策略。
急尖长苞冷杉(Abiesgeorgeivar.smithii)在西藏东南部主要分布于海拔3500~4300 m的高山地带,属阴性树种,是色季拉山乔木层植被群落的主要建群种,对藏东南林区水土保持、水源地养护以及区域生态平衡维护等具有不容忽视的作用[6],同时也担负着西藏生态安全屏障的主要作用。本研究以不同海拔梯度冷杉群落作为研究对象,选取植株最大高度、叶面积两个功能性状指标,研究了不同坡向、不同海拔冷杉群落木本植物的功能多样性,以了解色季拉山不同海拔梯度冷杉群落木本植物组成特征,以及生境条件对群落木本植物功能性状形成的作用与影响。
2 研究方法
2.1 研究区概况
研究样地位于西藏自治区林芝市色季拉山西藏林芝森林生态系统国家定位观测研究站附近,地理坐标29°38′02″~29°38′51.4″N,94°42′16.9″~94°43′13.3″E。分布于海拔3700~4300 m,属亚高山寒温性湿润气候,年均气温-0.73~1.8℃,6~9月份为雨季,雨量充沛,热量充足,年均降水量约1134 mm,年均相对湿度78.8%[7]。冷杉群落样地基本概况见表1。

表1 不同海拔冷杉群落样地基本概况
2.2 样地设置与调查
在色季拉山3700~4300 m海拔范围内,选取北坡及南坡两个不同坡向,以100 m海拔为间隔,共设置14个30 m×30 m冷杉群落样地。在生长季结束后,对样地内木本植物群落结构进行调查。每个物种选取5~10株长势良好的成年个体,测量其高度,并记录植株最大高度(HMAX)。从植株个体向阳面的冠层附近剪取至少3根比较完整的末端小枝,于当天每个植株选取至少20片完整的当年生成熟叶片或复叶中的小叶,用LI-3100C叶面积扫描仪扫描平展叶片的图像,得到叶面积(LA)数据(单位:cm2)。
3 结果与分析
3.1 群落木本植物物种组成
色季拉山不同海拔、不同坡向冷杉样地群落木本植物分布格局如表2所示,群落内乔木树种主要为急尖长苞冷杉,阳坡灌木树种主要有杯萼忍冬(Lonicerainconspicua)、冰川茶藨子(Ribesglaciale)、峨眉蔷薇(Rosaomeiensis)、柳叶忍冬(Loniceranigra)、西南花楸(Sorbusrehderiana)、小舌紫菀(Asteralbescens)、长刺茶藨子(Ribesalpestre)、紫斑杜鹃(Rhododendronstrigillosumvar.monosematum)、硬毛杜鹃(Rhododendronhirtipes)、红点杜鹃(Rhododendrontanastylumvar.lingzhiense)、蓝果忍冬(Loniceracaerulea)、雪山杜鹃(Rhododendronaganniphum)、金露梅(Potentillafruticosa)、林芝杜鹃(Rhododendronnyingchiense)等。阴坡灌木树种主要为糙皮桦(Betulautilis)、峨眉蔷薇、三花杜鹃(Rhododendrontriflorum)、硬毛杜鹃、直立悬钩子(Rubusstans)、杯萼忍冬、西南花楸、冰川茶藨子、陇塞忍冬(Loniceratangutica)、雪山杜鹃、红点杜鹃等。总体而言,色季拉山不同坡向冷杉群落木本植物种类较为丰富,阳坡树种数量要大于阴坡。

表2 海拔梯度不同坡向冷杉群落木本植物分布格局
3.2 群落木本植物物种数量分布特征
不同海拔、不同坡向样地群落木本植物物种数如图1所示。从海拔梯度来看,3700 m处不同坡向物种数最多,共12种;4300 m处物种数最少,为5种。阳坡木本植物表现为沿海拔上升,物种数呈现下降的趋势;阴坡物种数总体上也表现为随海拔上升而降低的趋势。但是在3800 m处,物种数量突然急剧下降。低海拔物种数总体大于高海拔。

图1 不同海拔样地群落木本植物物种数量特征
3.3 不同海拔木本植物LA分布特征
如图2所示,由于样地内植物在不同海拔梯度分布有差异且不连续,因此统计时选定至少在4个海拔上有分布的木本植物(下同)。阳坡不同海拔冷杉LA变化范围为0.36~0.59 cm2,最大值在4000 m海拔处,4200 m海拔LA最小;随海拔上升呈波动下降趋势。杯萼忍冬LA变化范围为1.61~3.90 cm2,样地内分布上限4100 m处LA最大,3700 m处最小,总体呈随海拔上升呈增长趋势。峨眉蔷薇LA变化范围为8.78~17.25 cm2,最小值在3900 m,最大值同样位于样地分布最高海拔4200 m处,总体变化趋势为随海拔上升呈波动增长。柳叶忍冬LA变化范围为9.40~14.81 cm2,3700 m处LA最小,样地分布上限4000 m处最大,LA随海拔上升呈逐渐递增趋势。小舌紫菀LA变化范围为14.58~18.23 cm2,与前几种相反,样地分布上限4000 m处LA最小,3700 m处最大,LA随海拔增加呈逐级递减趋势。紫斑杜鹃LA变化范围为21.86~29.09 cm2,3900 m处值最小,样地内分布上限4200 m处LA最大,变化趋势为随海拔增加先降后升。
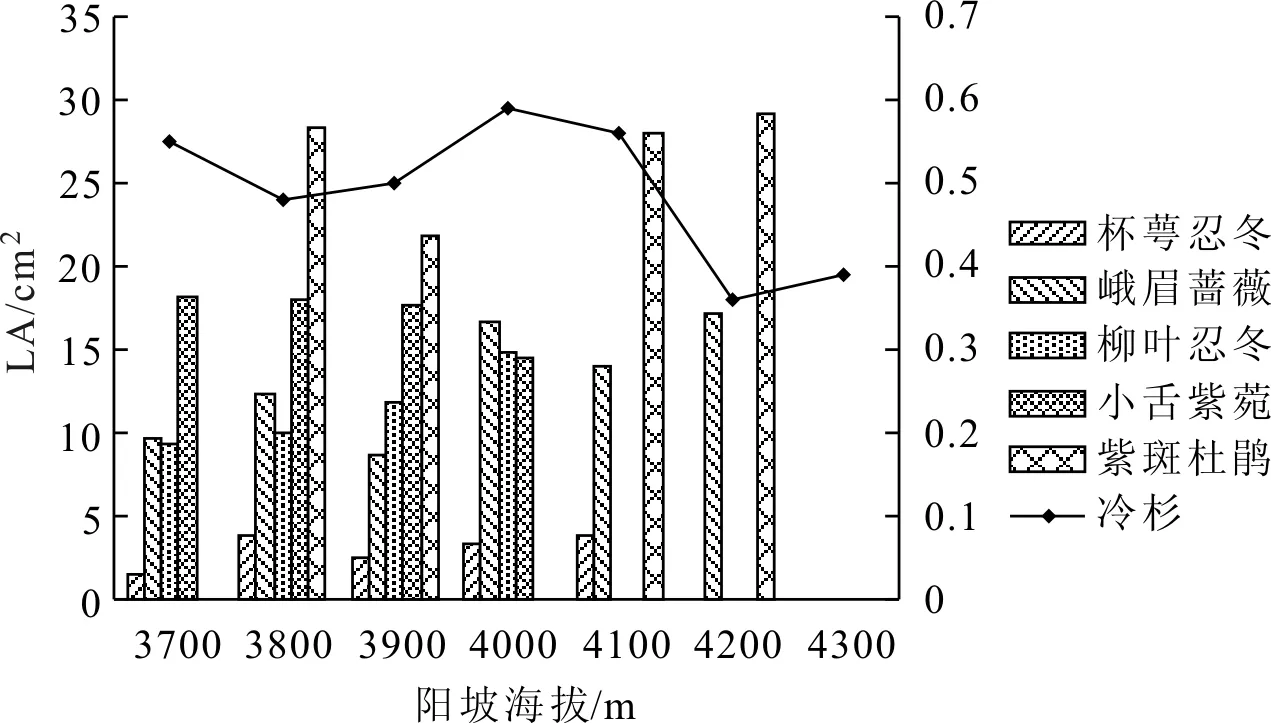
图2 阳坡叶面积垂直分布特征
如图3所示,阴坡不同海拔冷杉LA变化范围为0.34~0.60 cm2,最大值在3700 m处,4300 m海拔LA最小;随海拔增加呈波动下降趋势。峨眉蔷薇LA变化范围为13.06~20.14 cm2,3800 m值最大,3900 m处值最小,随海拔无明显变化规律。硬毛杜鹃LA变化范围为27.58~34.14 cm2,最大值在3900 m处,最小值3700 m处,总体随海拔上升有所增大。西南花楸LA变化范围为21.09~42.93 cm2,样地分布最高海拔4200 m处LA最大,最低海拔3900 m处LA最小,LA随海拔上升呈逐渐递增趋势。
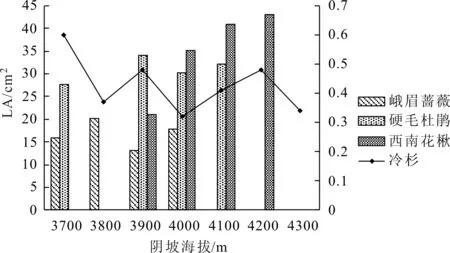
图3 阴坡叶面积垂直分布特征
3.4 不同海拔木本植物HMAX分布特征
如图4所示,阳坡不同海拔冷杉HMAX变化范围约为20.0~41.5m,样地分布最低海拔3700 m处HMAX最高,最低海拔4300 m处HMAX最小,HMAX随海拔上升呈递减趋势。杯萼忍冬变化范围为2.4~5.3 m,样地内分布上限4100 m处HMAX最大,3800 m处最小,随海拔上升表现为先降后增趋势。峨眉蔷薇变化范围为2.5~4.8 m,最小值分布在4200 m,最大值位于样地分布最低海拔3700 m处,总体变化趋势为随海拔上升呈波动下降。柳叶忍冬变化范围为3.8~5.1 m,4000 m处HMAX最小,3700 m处高度最大,随海拔上升呈波动递减趋势。小舌紫菀HMAX变化范围为0.8~1.5 m,4000 m处HMAX最小,4100 m处最大,HMAX随海拔增加呈先降后升趋势。紫斑杜鹃变化范围为2.5~8.5 m,4100 m处值最小,3900 m处HMAX最大,其随海拔增加变化趋势不明显。

图4 阳坡最大株高垂直分布特征
如图5所示,阴坡不同海拔冷杉HMAX变化范围约为11.2~37.5 m,最大值在3900 m处,4300 m海拔HMAX最小,随海拔增加呈先增后减降趋势。峨眉蔷薇变化范围为1.6~2.5 m,3700 m值最大,3900 m处值最小,随海拔增加呈先降后增趋势。硬毛杜鹃变化范围为2.2~6.0 m,最大值在样地分布最高海拔4100 m处,最小值4000 m处,随海拔增加HMAX无明显变化趋势。西南花楸HMAX变化范围为2.9~6.9 m,4100 m处HMAX最大,样地分布最低海拔3900 m处最小,HMAX总体随海拔上升呈增长趋势。

图5 阴坡最大株高垂直分布特征
4 讨论
从数量分布格局来看,群落内木本植物主要表现为沿海拔梯度上升,物种数呈逐渐下降趋势。由于环境梯度因素,尤其是气温,在垂直尺度上具有较强的空间差异性[8],因此这种因素有助于在群落间产生环境过滤作用,低海拔较好的水热条件导致群落木本植物物种数高于高海拔的总体特征。坡度是地形影响因素之一,它可以通过影响土壤养分、水分等环境因素,从而决定群落物种的组成和分布[9]。3700~3900 m样地阴坡物种数>阳坡,可能是由于坡度因素导致。3700~3900 m海拔处阳坡坡度较阴坡小(表1),受雨水冲刷强度低,土壤有机质含量较阳坡更高,因此坡度小更利于多种植物的定植[10],从而物种数量更高。4000~4300 m阳坡坡度虽然较阴坡更大,但是海拔较高冷杉林也越稀疏,郁闭度更低,因而利于林下其他木本植物定植生长,所以这些群落阳坡阴坡物种数大致相等。
从LA分布特征来看,不同坡向冷杉LA均沿海拔上升呈波动下降趋势。主要是由于在3700~4300 m的垂直海拔梯度上,水热条件差异较大,低海拔水热条件较好,作为群落优势种的冷杉因而具有较高的LA。这与向琳等[11]对井冈山鹿角杜鹃群落灌木层植物叶功能性状的研究一致,也与田俊霞等[12]对温带针阔混交林叶片性状垂直高度上的分布规律一致。阳坡杯萼忍冬、峨眉蔷薇、柳叶忍冬、紫斑杜鹃LA表现为随海拔上升总体呈增加趋势。这是由于海拔上升群落内冷杉林郁闭度降低,光照条件较好,林下植物光合作用增强,从而LA更大,这与Zhang等[13]的研究结果相似。阴坡峨眉蔷薇、硬毛杜鹃、西南花楸LA总体上同阳坡一致,均表现为在其适生环境梯度上随海拔增加,LA也增加的趋势。阳坡小舌紫菀LA随海拔增加表现为逐级降低。样地内小舌紫菀LA最大为18.23 cm2,且总体比上述物种LA更大。由于海拔越高,低温条件对LA更大的植物生理影响越大,因而导致小舌紫菀随海拔增加LA越小。
从植株HMAX分布特征来看,冷杉阳坡HMAX介于20~41.5 m,阴坡范围为11.2~37.5 m,总体上阳坡HMAX>阴坡,且不同坡向沿海拔梯度上升均主要表现为下降的规律。这是由于冷杉属于耐阴性树种[14],适于生长在光照强度低、阴凉潮湿的生境中。同时,由于低海拔在土温、水分等方面的良好优势,因此不同坡向HMAX均表现为低海拔>高海拔。阳坡峨眉蔷薇、柳叶忍冬、紫斑杜鹃及阴坡峨眉蔷薇HMAX同冷杉变化趋势一致,其原因同样与水热条件的垂直变化有关[15]。阳坡杯萼忍冬、小舌紫菀以及阴坡硬毛杜鹃、西南花楸HMAX随海拔上升总体呈上升趋势,这是由于海拔越高,林分郁闭度越低,林下植物获得充足光照的可能性更大,从而促进了植物生长。
5 结论
色季拉山不同海拔梯度、不同坡向冷杉群落木本植物种类较为丰富,阳坡树种数量>阴坡;低海拔物种数总体>高海拔。
不同坡向冷杉叶面积均沿海拔梯度上升总体呈波动下降规律。阳坡主要林下木本植物叶面积表现为随海拔上升总体呈增加趋势;阴坡植物叶面积变化趋势总体上同阳坡一致。
不同坡向冷杉最大高度总体沿海拔梯度上升逐渐降低。阳坡木本植物最大高度变化规律同冷杉变化趋势一致;阴坡植物随海拔增长总体上最大高度也增加。由此可见,水热条件等环境异质性深刻塑造了沿海拔梯度的森林群落构建格局及其功能多样性特征。
猜你喜欢
世界热带农业信息(2022年8期)2022-07-19
山地农业生物学报(2022年3期)2022-05-13
土壤学报(2022年1期)2022-03-08
种子科技(2021年8期)2021-07-11
学校教育研究(2020年7期)2020-04-09
华东师范大学学报(自然科学版)(2019年3期)2019-06-24
福建基础教育研究(2019年3期)2019-05-28
西部资源(2018年1期)2018-11-01
中学生理科应试(2017年6期)2017-09-27
中学生数理化·高三版(2016年9期)2016-05-14
