
长链非编码RNA ZFAS1在结直肠癌中的表达及生物学功能
2022-05-28 05:57武丽娜彭小忠鲁重美
生物技术进展 2022年3期
武丽娜, 彭小忠, 鲁重美*
1.中国医学科学院北京协和医学院北京协和医院消化内科,北京100730;2.中国医学科学院北京协和医学院基础医学研究所,医学分子生物学国家重点实验室,北京100730
结直肠癌是威胁人类身体健康的高发肿瘤,据统计,我国结直肠癌死亡率在所有肿瘤中居第5 位,该病5 年生存率为66.1%,但出现远处转移时,患者5 年生存率会降至12.5%[1]。结肠腺瘤-腺癌序列最早是由病理学家Basil 发现并定义的,此后该序列各步骤发生发展的分子机制被逐渐完善[2],并通过分子遗传学研究揭示了一些新的癌基因及抑癌基因参与了结直肠癌的发生发展[3]。人类基因组大约75%的基因可转录为RNA,其中仅有约2%的基因与编码蛋白相关,其余均为非编码RNA[4]。长链非编码RNA(long noncoding RNA,lncRNA)是一类长度超过200 nt、无蛋白质编码功能的RNA 分子,其可以在转录水平、转录后水平及表观遗传水平发挥作用[4-5],同时与部分肿瘤的发生发展有关[6-7]。LncRNA 锌指核转录因子反义链1(zinc finger nuclear transcription factor antisense RNA 1,ZFAS1)位于20q13,是锌指核转录因子1(zinc finger nuclear transcription factor,Xbox binding 1-type containing 1,ZNFX1)的 反 义RNA,同时也是3 个C/D 盒小核仁RNA(C/D-box snoRNAs,SNORDs),即SNORD12C、SNORD12B及SNORD12 的载体。ZFAS1 包括10 种转录本,长度为504~1 075 bp。ZFAS1 的全长尚不清楚,且目前也鲜见对ZFAS1 进行cDNA 末端快速扩增技术(rapid amplification of cDNA ends,RACE)实验的报道。本研究旨在探讨ZFAS1 在结直肠癌中的表达及生物学功能,并以ZNFX1 为切入点,以期明确ZFAS1 在结直肠癌中的作用机制。
1 材料与方法
1.1 研究对象
选取2014 年10 月—2015 年7 月于北京协和医院进行结直肠癌手术,且手术病理结果证实为结直肠癌的患者67 例作为研究对象,其中男37例,平均年龄63 岁;女30 例,平均年龄65 岁。统计所有患者的临床资料及病理资料,并将所有患者结直肠癌标本及配对正常结直肠组织浸入RNA laterTM(Ambion 公司)中,置于−20 ℃冰箱保存。所有患者均对本研究知情并自愿签署知情同意书,本研究已通过北京协和医院医学伦理委员会审核批准。
1.2 材料
人结肠癌细胞系HT-29、LoVo、SW480、HCT-116、RKO、HCT-8、HCT-15、DLD1及人结肠上皮细胞系CCD841CoN 均购自北京协和医学院基础医学研究所细胞中心,且均源于美国模式培养物集存库(https://www.atcc.org/)。
1.3 方法
1.3.1 实时荧光定量PCR(real-time PCR)测定目的基因在结肠癌细胞系中的表达水平 所有组织及细胞均由总RNA 抽提试剂TRIzol(Invitrogen 公司)裂解并提取总RNA,按照逆转录试剂盒(Roche 公司)说明书进行反转录,按照SYBR Premix Ex Taq试剂盒(TaKaRa公司)说明书进行realtime PCR检测,引物序列见表1。

表1 ZFAS1和ZNFX1的real-time PCR引物Table 1 The real-time PCR primers of ZFAS1 and ZNFX1
1.3.2 Western blot 实验测定ZNFX1 在结肠癌细胞系中的表达 裂解结肠癌细胞,根据BCA 蛋白试剂盒(Abcam 公司)说明书检测蛋白质浓度。蛋白经8%SDS-PAGE 胶分离、转膜、封闭,分别经一抗(抗ZNFX1抗体或抗β-actin抗体)及二抗孵育,采用增强化学发光法(enhanced chemiluminescence,ECL)显影。利用Image J 软件计算各蛋白条带吸光度与β-actin 吸光度的比值,并进行半定量分析。
1.3.3 通过转染小干扰RNA(small interfering RNA,siRNA)敲低目的基因 根据INTERFERinTM(Polyplus 公司)说明书将siRNA(30 pmol·mL−1)转染至LoVo、SW480、HCT-116细胞。针对ZFAS1基因的2个siRNA序列见表2。

表2 siRNA序列Table 2 The sequences of siRNA
1.3.4 通过转染表达载体质粒过表达目的基因 构建ZNFX1 CDS 区第1~5 757 位的克隆质粒并命名为ZNFX1-1-5757bp,该表达载体质粒包括ZN-FX1 的全长;构建ZNFX1 CDS 区第1~3 646 位的克隆质粒并命名为ZNFX1-1-3646bp,该表达载体质粒包括ZNFX1 的两个AAA 结构域。根据Lipofectamine 2000(Invitrogen 公司)说明书将这两个表达载体质粒转染至LoVo、SW480、HCT-116 细胞,从而过表达克隆质粒所携带的基因片段。
1.3.5 细胞活力试验测定结肠癌细胞的增殖能力在96 孔板中将各组细胞以3×103个细胞·孔−1铺板,根据细胞增殖实验试剂盒(Promega 公司)说明书,将四氮唑化合物[3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium,MTS]及电子偶联剂(phenazine methosulfate,PMS)按体积比20∶1 配制MTS/PMS 溶液,将MTS/PMS 与细胞培养基按体积比为1∶5 加入培养基中,继续在37℃、5% CO2孵箱培养4 h。以630 nm 波长处的吸光度值作为校正,分别取5 个时间点(转染后0、24、48、72、96 h)在490 nm 波长处检测吸光度,设3 次生物学重复。
1.3.6 平板克隆形成试验测定结肠癌细胞的增殖能力 在转染后24 h,在6 孔板中将各组细胞以3×103个细胞·孔−1铺板,置于37 ℃、5%CO2及饱和湿度(95%)环境下,静置培养10~14 d,待各孔中出现肉眼可见克隆时终止培养,并采用4%多聚甲醛固定,2.5%结晶紫染色,计数每孔克隆数目,设3次生物学重复。
1.3.7 Transwell迁移试验测定结肠癌细胞的迁移能力 以1×105个细胞·孔−1将各组细胞接种于Transwell上层小室中,小室中加入无血清培养基,在下层小室中加入含有10%胎牛血清的培养基。在37 ℃、5% CO2孵箱孵育24~48 h,并采用4%多聚甲醛固定,2.5%结晶紫染色,在M205 体视显微镜(Leica 公司)下统计迁移到下层小室的细胞数,设3次生物学重复。
1.4 统计学分析
采用SPSS 20.0 统计软件进行数据分析,计量资料以平均数±标准差(x±s)表示,两独立样本间比较采用t检验,多组样本间比较采用One-Way ANOVA 检验,两独立样本的四格表资料采用Chi-square 检验;以P<0.05 为差异有统计学意义。
2 结果与分析
2.1 ZFAS1 在结直肠癌组织及结肠癌细胞系中的表达水平
如图1 所示,ZFAS1 在结肠癌细胞系(DLD1、HCT-116、LoVo、HCT-8、HT-29 及RKO)中的相对表达量显著高于正常结肠上皮细胞系(CCD841 CoN),差异有统计学意义(P<0.05)。ZFAS1 在结直肠癌组织中的相对表达量(0.56±0.09)显著高于癌旁正常组织(0.23±0.04),差异有统计学意义(P<0.05)。结果表明,ZFAS1 在结直肠癌组织及结肠癌细胞系中高表达。
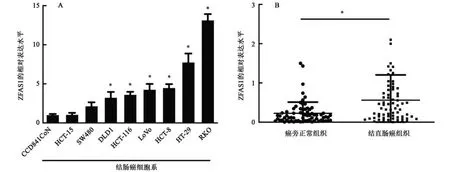
图1 ZFAS1在结肠癌细胞系和结直肠癌组织中的表达水平Fig.1 The expression levels of ZFAS1 in colorectal cancer cell lines and colorectal cancer tissues
2.2 ZFAS1 在结直肠癌中表达水平与临床病理学资料的相关性
ZFAS1在结直肠癌组织中相对表达量的中位数为0.35,并以此作为分界值将67 例患者分为高表达组(≥0.35)及低表达组(<0.35)。由表3可知,高表达组与低表达组在性别、年龄、血癌胚抗原水平、肿瘤部位、肿瘤大小、分化程度、肿瘤浸润深度、淋巴结转移、远处转移及TNM 分期比较差异无统计学意义(P>0.05)。结果表明,ZFAS1 在结直肠癌组织中高表达或低表达与结直肠癌患者的临床病理学情况无显著相关性。

表3 ZFAS1的表达与结直肠癌患者的临床病理资料的相关性Table 3 Correlation between ZFAS1 and clinicopathological data of colorectal cancer patients n(%)
2.3 敲低ZFAS1 对结肠癌细胞的生长及迁移的影响
选取3 株应用广泛,细胞培养难度低结肠癌细胞系HCT-116、LoVo 及SW480 进行后续的生物学功能试验。检测ZFAS1 siRNAs 转染结肠癌细胞系的敲低效果,由图2 可知,与control siRNA 相比,ZFAS1 siRNA1 及ZFAS1 siRNA2 均可显著下调ZFAS1的表达量(P<0.05),且ZFAS1 siRNA1的作用更明显,因此,选用ZFAS1 siRNA1 进行后续细胞功能学试验。敲低ZFAS1 后,结肠癌细胞系HCT-116、LoVo 及SW480 的克隆形成数量均少于control siRNA。敲低ZFAS1 后,结肠癌细胞系HCT-116、LoVo 及SW480 的活细胞数量均显著少于control siRNA(P<0.05)。敲低ZFAS1 后,结肠癌细胞系HCT-116、LoVo 及SW480 穿过Transwell微孔滤膜的细胞数量均少于control siRNA。结果表明,敲低ZFAS1,结肠癌细胞的生长能力和迁移能力均下降,提示ZFAS1 可以促进结肠癌细胞的生长和迁移。
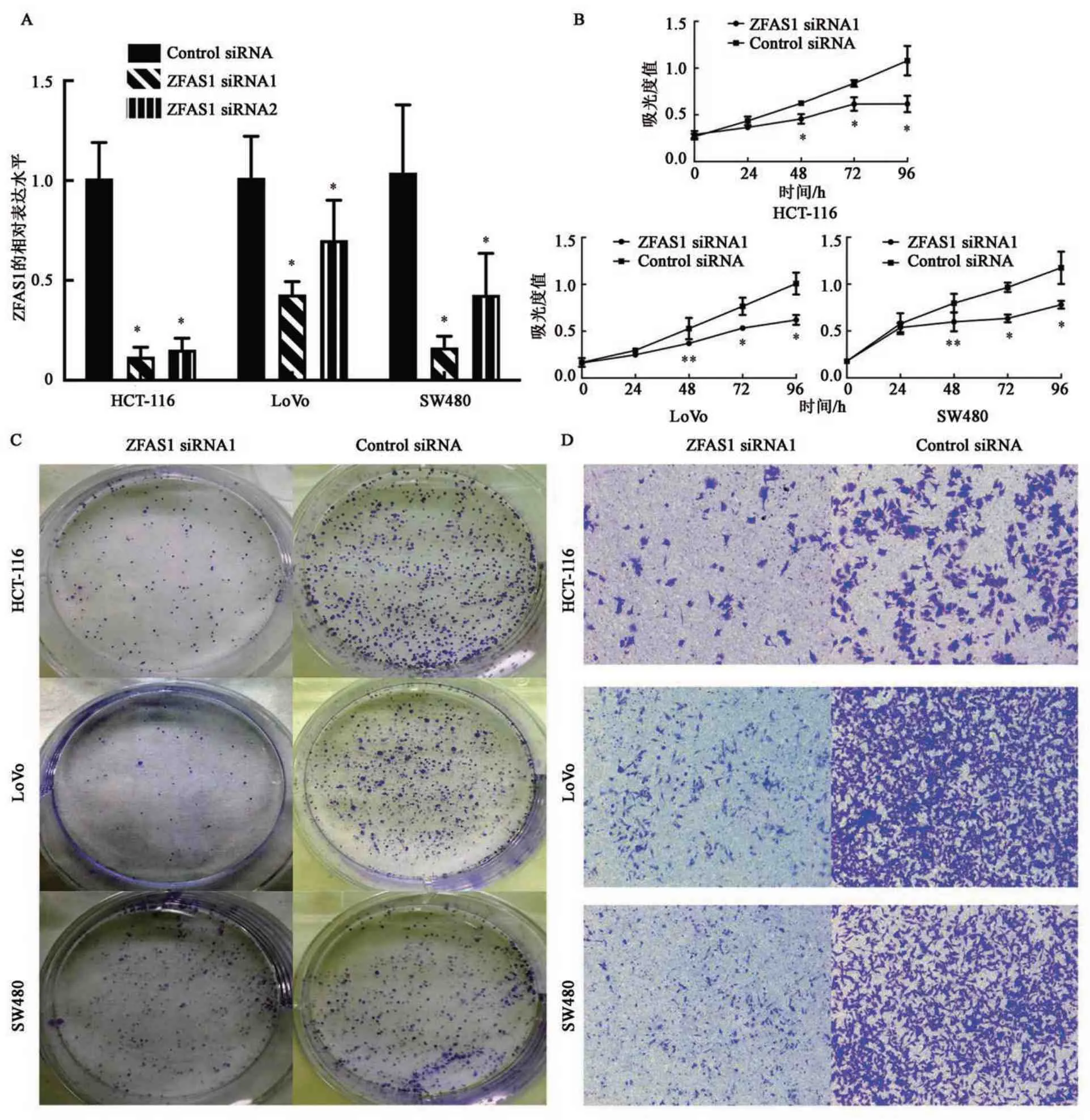
图2 敲低ZFAS1对结肠癌细胞的生长及迁移的影响Fig.2 Knockdown effects of ZFAS1 on the growth and migration of colorectal cancer cells
2.4 敲低ZFAS1对ZNFX1表达的影响
由图3 可知,ZNFX1 在结肠癌细胞系(LoVo、DLD1、HCT-116、SW480、HCT-8)中的相对表达量均低于正常结肠细胞系CCD841CoN。敲低ZFAS1 后,与ZFAS1 相比,ZNFX1 表达水平上调(P<0.05)。结果表明,ZNFX1 在结肠癌细胞系中低表达,ZFAS1可以负调控ZNFX1的表达水平。

图3 ZFAS1与ZNFX1表达量的相关性Fig.3 The relationship between the expression level of ZFAS1 and ZNFX1
2.5 过表达ZNFX1 对结肠癌细胞生长及迁移的影响
目前对于ZNFX1 编码蛋白的晶体结构尚无解析结果,本研究根据蛋白质结构预测在线软件(http://smart. embl-heidelberg.de/)对ZNFX1 进行结构域预测:ZNFX1 的肽链从N 端到C 端依次是由2 个AAA 结构域及3 个锌指结构域组成(图4A)。将两种表达载体质粒(ZNFX1-1-5757bp和ZNFX1-1-3646bp)分别转染结肠癌细胞,检测过表达效果,结果表明,转染ZNFX1-1-5757bp 的结肠癌细胞中ZNFX1 的表达量高于pcDNA-NC,转染ZNFX1-1-3646bp的结肠癌细胞在130~180 kD分子量之间出现一条新的条带,即为构建的ZNFX1-1-3646bp 克隆质粒所表达出的蛋白(图4B)。过表达ZNFX1-1-5757bp 及ZNFX1-1-3646bp 后,结肠癌细胞系HCT-116、LoVo 及SW480 的活细胞数量显著少于pcDNA-NC(P<0.05,图4C),且结肠癌细胞系HCT-116、LoVo 及SW480 的克隆形成数量少于pcDNA-NC(图4D)。同时,结肠癌细胞系HCT-116、LoVo 及SW480 穿过Transwell 微孔滤膜的细胞数量少于pcDNA-NC(图4E)。结果表明,ZNFX1可以抑制结肠癌细胞的生长及迁移,过表达ZNFX1,结肠癌细胞的生长及迁移能力下降,并且ZNFX1的AAA结构域区段可以独立发挥该功能。


图4 过表达ZNFX1对结肠癌细胞的生长及迁移的影响Fig.4 Proliferation and migration of CRC cells after ZNFX1 overexpression
3 讨论
研究表明ZFAS1 在不同肿瘤组织中的表达情况存在差异[8],如Askarian-amiri 等[9]研究发现,ZFAS1 在正常乳腺中高表达,在乳腺侵袭性导管腺癌中表达下调;大量研究发现,ZFAS1 在胃癌[10]、肾透明细胞癌[11]、骨肉瘤[12]中均为高表达。本研究发现ZFAS1 在结直肠癌组织及结肠癌细胞系中表达升高,并且可以促进结肠癌细胞的生长及迁移,证实ZFAS1 在结直肠癌中具有原癌基因的潜质,这与既往ZFAS1 在结直肠癌中的相关研究结果相符[13]。
ZFAS1在结肠癌细胞中的功能机制多样,既往研究主要从以下几个方面进行探索:①ZFAS1 作为内源性竞争性RNA(competing endogenous RNAs,ceRNAs)与miRNA 结合,作为“miRNA 海绵”阻断miRNA 介导的抑癌功能。在宫颈癌中,ZFAS1可与miR-190a-3p结合阻断miR-190a-3p介导的抑癌功能[14]。在结直肠癌中,ZFAS1作为ceRNAs 可与miR-144[15]、miR-7-5p[16]、miR-150-5p[17]、miR-484[18]等miRNA 结合,促进结直肠癌的增殖及转移。②ZFAS1是3个SNORDs的载体,既往研究提示,lncRNA 可以通过snoRNA 起到促进肿瘤增殖及转移的作用,如GAS5基因在结肠癌中的抑癌作用可能与由GAS5衍生的snoRNA相关,且研究表明在DNA损伤信号通路中,p53诱导GAS5snoRNA表达,从而起到促进凋亡并减慢细胞周期的作用[19]。ZFAS1 在结直肠癌中的作用是否与SNORDs 相关尚并不清楚,有待进一步研究。③ZFAS1是ZNFX1的反义RNA,二者“头对头”转录,ZFAS1及ZNFX1 在小鼠肺、乳腺、肾脏、脑、脾、心、肝、胚胎等组织中均有表达[8],且在大多数组织中两者的表达水平存在相关性。在侵袭性导管腺癌中ZFAS1 表达下调,而ZNFX1 表达上调[9]。在结肠癌中ZFAS1的功能是否与ZNFX1相关尚不明确。
本研究发现,敲低ZFAS1 后,ZNFX1 表达上调,表明ZFAS1 与ZNFX1 在结肠癌细胞系中的表达水平具有相关性,ZFAS1 能够潜在负调控ZNFX1 的表达。ZNFX1 在结肠癌细胞系中表达下调,表明ZNFX1 在结肠癌细胞及正常结肠细胞中存在差异性表达。细胞功能学试验证明,ZNFX1可以抑制结肠癌细胞的生长及迁移,提示ZFAS1可能通过抑制ZNFX1 的表达从而起到促进结肠癌细胞增殖及转移的作用。此外ZNFX1 的肽链是由2 个AAA 结构域及3 个锌指结构域组成。AAA 结构域与一系列细胞活性相关,包括膜融合、蛋白质水解及DNA 复制等[20-21]。锌指结构域属于相对较小的蛋白模体,通过指样结构域识别DNA、RNA、蛋白质及脂质底物等[22]。为确定ZNFX1 哪个区段在结直肠癌发生发展中起到作用,本研究采用截短法,通过转染不同结构域区段的质粒进行生物学功能试验,证明ZNFX1的AAA结构域区段可以独立发挥抑制结肠癌细胞生长及迁移的功能。
综上所述,本研究结果表明,ZFAS1在结直肠癌组织及结肠癌细胞系中表达上调,可以促进结肠癌细胞的生长及迁移,即ZFAS1 具有原癌基因的潜质,且ZFAS1 可能通过抑制其潜在靶基因ZNFX1 的表达从而起到促进结肠癌细胞增殖及转移的作用。但本研究未发现ZFAS1 的高表达或低表达与结直肠癌患者的临床病理学资料之间有统计学意义,考虑可能与纳入的患者例数较少有关,未来需进一步扩大样本量评估二者的相关性。另外,本研究发现ZFAS1 可能通过抑制ZNFX1的表达从而起到促进结肠癌细胞增殖及转移的作用,但具体的分子机制尚不清楚,未来需进一步完善分子生物学机制研究。
猜你喜欢
湖北农业科学(2022年11期)2022-07-18
实用肿瘤学杂志(2020年4期)2020-12-08
天津医科大学学报(2019年6期)2019-08-13
腹腔镜外科杂志(2016年10期)2016-06-01
腹腔镜外科杂志(2016年10期)2016-06-01
医学研究杂志(2015年9期)2015-07-01
癌变·畸变·突变(2015年4期)2015-02-27
中国老年学杂志(2015年9期)2015-01-31
中华介入放射学电子杂志(2014年1期)2014-02-02
中国医学科学院学报(2012年3期)2012-03-25
