
远红光有效促进罗勒植株的生物量和叶片花青素、总酚及类黄酮的积累
2022-05-30 13:36窦海杰GenhuaNiuMengmengGuJosephMasabni
中国蔬菜 2022年5期
窦海杰 Genhua Niu Mengmeng Gu Joseph Masabni
(1 北京一轻研究院有限公司,智能化室内植物种植研发平台,北京 101111;2Texas A &M AgriLife Research,Dallas,TX,USA 75252;3Department of Horticultural Sciences,Texas A &M AgriLife Extension Service,College Station,TX,USA 77843)
光合有效辐射(PAR,400~700 nm)不仅为植物的光合作用提供能量,而且可作为一种信号调控因子调控作物的生长发育(Snowdenet al,2016;Douet al.,2017,2019;Son et al.,2017)。光合有效辐射波段外的波长,如紫外辐射(UR,280~399 nm)和远红光辐射(FR,700~780 nm)同样可调控作物的生长发育(Casal,2013;Wargent &Jordan,2013;Ballaré,2014)。近年来研究表明,远红光可调控植株的光合作用、光形态建成及植株的部分次生代谢产物含量(Yang et al.,2013;Demotes-Mainard et al.,2016;Zhen &van Iersel,2017;Meng et al.,2019;Zhen &Bugbee,2020)。研究发现光合有效辐射与远红光辐射能够协同提高植株的净光合速率,且其量子产额大于光合有效辐射及远红光辐射单独照射的总和,此现象被称为增益效应或爱默生效应(Emerson et al.,1957;Emerson &Rabinowitch,1960)。增益效应的发现表明植株中存在2 个光系统,即光系统I(PSI)和光系统II(PSII),分别由远红光辐射和光合有效辐射优先激活(Emerson &Rabinowitch,1960;Myers,1971;Allen,2003),且2 个光系统协同进行光合作用的电子传递。单独的光合有效辐射或远红光辐射使得PSI 和PSII 处于失衡状态,光量子产额较低。
人工光型植物工厂一般使用仅包含光合有效辐射波段的人工光源作为植株光合作用的唯一光源。在光合有效辐射中加入远红光能够有效驱动PSI 的循环电子传递,缓解电子受体端的限制,提高光系统的光量子效率(Kono et al.,2019)。研究表明,生长在红蓝光(RGB)和白光(BGRFR)下的叶用莴苣(生菜)植株,添加远红光后植株PSII 的光量子效率和植株净光合速率快速增加(Zhen &van Iersel,2017)。在光合有效辐射的基础上添加总光强比例约40%的远红光对植株冠层净光合速率的增加值与添加相同强度的光合有效辐射的增加值相同(Zhen &Bugbee,2020)。在光合有效辐射强度为200 μmol·m·s的条件下,添加50 μmol·m·s的远红光可使PSI 和PSII 达到平衡,但继续增加远红光的光照强度对PSII 光量子效率的增加无促进作用(Zhen &van Iersel,2017)。
远红光不仅影响植株的光合作用效率,同时还调控植株的光形态建成(Trupkinet al.,2014;Park &Runkle,2017)。远红光通过光敏色素phyB调控光敏色素相互作用因子(PIFs),激活植株生长素合成基因,诱导植株的庇荫反应(Trupkin et al.,2014)。若红光∶远红光(R∶FR)>1,光敏色素phyB 由红光吸收态(P,λ=660 nm)转变为远红光吸收态(P,λ=730 nm),进而通过PIF7 调控路径诱导生长素的合成及植株的庇荫反应(Quail,2002;Liet al.,2012)。在红蓝光中增加远红光使得叶用莴苣和罗勒叶面积增大,植株鲜质量增加,但植株叶绿素含量降低。结果表明,R∶FR 值较低时,植株茎秆和叶柄伸长、叶片倾角向上、侧枝减少,增加了植株对光合有效辐射的捕获(Meng &Runkle,2017)。
综上可知,在光合有效辐射的基础上添加远红光能够促进植株的光合作用,调控光形态建成,增加植株生物量积累。因此,有研究建议将光合有效辐射的波段范围扩大至远红光波段(Franklin,2008;Zhen &Bugbee,2020)。但是,在相同的总光照强度下,用远红光代替部分光合有效辐射对植株生长发育的作用机理仍不明确,比例不当有可能造成植株光合有效辐射捕获量降低,对植株生物量积累产生抑制作用,以及减弱远红光对植株光合、生物量积累的促进作用。因此,探究远红光代替部分光合有效辐射对植株生长发育的影响,对人工光型植物工厂作物栽培的光环境调控具有重要意义。
罗勒()为唇形科罗勒属药食兼用芳香植物,别名九层塔、香花子、兰香等。植株含有丰富的芳香油及花青素、总酚和类黄酮等抗氧化物,可调配化妆品、香料、日用品和医药保健等产品(Sun et al.,2021)。本试验选取绿叶罗勒为研究对象,利用5 种不同的人工光源在植物工厂内开展远红光代替光合有效辐射对植株光合、形态、产量和部分次生代谢产物含量影响的研究,以期为人工光型植物工厂的光环境调控提供理论依据和实践支撑。
1 材料与方法
1.1 试验材料及栽培方法
于2019 年在美国德州农工大学埃尔帕索试验基地开展了远红光对罗勒植株生物量和叶片花青素、总酚及类黄酮积累影响的试验,以绿叶罗勒(Improved Genovese Compact,种子购于Johnny’s Selected Seeds)为试材,在温室内进行播种育苗,温室内的温度(昼/夜)为(28.3 ± 3.0)℃/(23.7 ± 1.4)℃,相对湿度(昼/夜)为(40 % ± 13%)/(56% ± 10%)。播种3 周后(幼苗展开3 片真叶时),将幼苗移栽至人工光型植物工厂实验室(35 m),采用基质栽培于口径约10 cm的方形苗钵中,栽培密度为79 株·m,每处理12株。按需浇灌营养液。营养液的EC 值为(2.01 ±0.06)dS·m,pH 值为5.98 ± 0.03。植株冠层温度(昼/夜)为(23.5 ± 0.5)℃/(22.0 ± 0.1)℃。移栽后进行不同光照处理,19 d 后统一采收,并进行相关指标的测定。
1.2 试验处理方法
设计5 种不同的人工光源,分为两组共10 个处理(表1)。第1 组为不含远红光(或远红光含量极低,忽略不计)的5 种人工光源处理,分别为红蓝光LED1(RB)、红蓝光LED2(RB)、红蓝光LED3(RB)、白色LED(RGB)和白色荧光灯(RGB)。第2 组即在保持总光量子通量密度(TPFD)不变的情况下,用远红光代替5 种人工光源的部分光合有效辐射的5 个处理,分别为红蓝光LED1/FR(RBFR)、红蓝光LED2/FR(RBFR)、红蓝光LED3/FR(RBFR)、白色LED/FR(RGBFR)和白色荧光灯/FR(RGBFR)。各处理的TPFD 见表1,光照周期均为16 h/8 h(昼/夜)。TPFD 和光合光量子通量密度(PPFD)为灯下15 cm 的测定值,根据各波段波长对植株光合作用的有效性加权获得各处理的有效光量子通量密度(YPFD)(表1)。
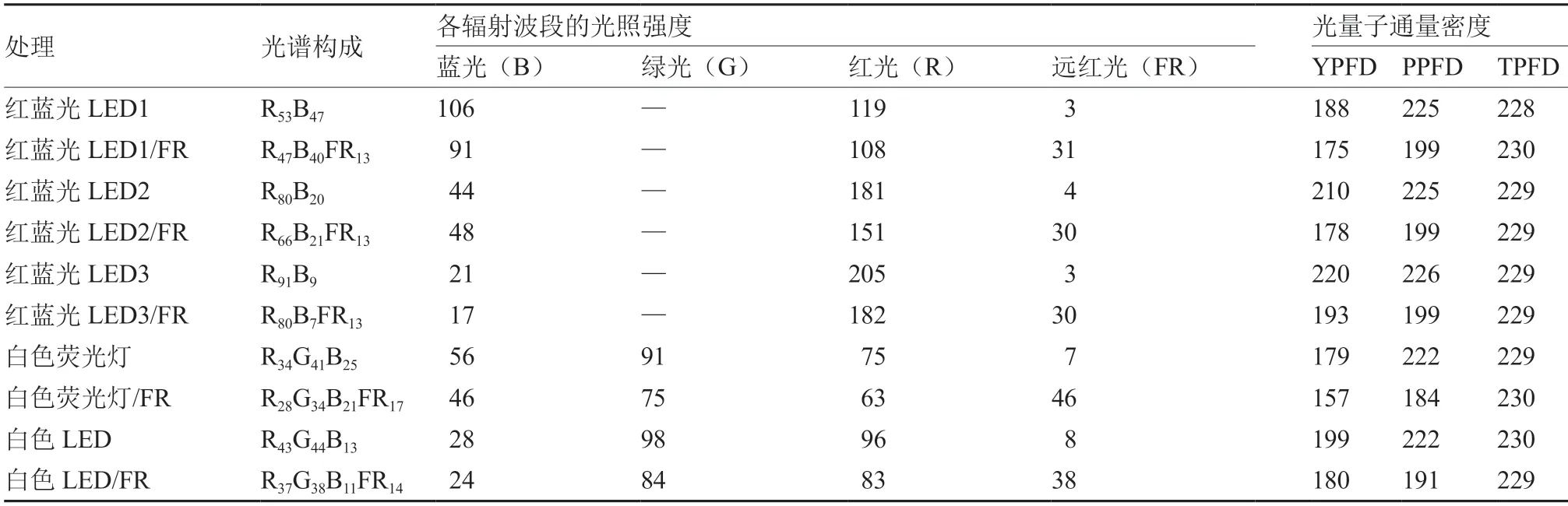
表1 各处理不同辐射波段的光照强度和光量子通量密度 μmol·m-2·s-1
1.3 项目测定
1.3.1 生长发育指标的测定 每个处理随机选取5株测定植株的株高、开展度、节间数、叶面积以及地上部、地下部的鲜质量和干质量,其中地上部分别测定了叶片及茎秆的鲜质量和干质量。叶面积使用叶面积仪(LI-3100,LI-COR)进行测定。将植株地上部置于80 ℃烘箱(Grieve)烘干至恒重,测定干质量。
1.3.2 光合速率及叶绿素含量的测定 每个处理随机选取5 株进行光合速率和叶绿素含量的测定。光合速率利用便携式光合测定仪(CIRAS-3,Portable Photosynthesis Systems International)的标准叶室(PLC3)进行测定,叶室内的光照强度、相对湿度、CO浓度分别为800 μmol·m·s、50%、390 μmol ·mol。叶片的叶绿素相对含量用便携式叶绿素测定仪SPAD-502(Konica-Minolta cooperation,Ltd.)进行测定,用SPAD 值表示;叶绿素a+b 含量采用浸提法进行测定(Porra et al,1989)。
1.3.3 花青素、总酚、类黄酮及抗氧化活性的测定 每个处理随机选取5 株测定植株叶片的花青素、总酚、类黄酮的含量和抗氧化活性。取新鲜的植物叶片于-80 ℃保存备用。测定时,称取约2 g叶片进行研磨,然后加入酸化甲醇溶液浸提,在黑暗中冷藏静置12~16 h 后离心,吸取上清液用于以上各项指标的测定(Xu &Mou,2016)。花青素含量采用分光光度法测定,测定结果以氰化物-3-葡萄糖苷当量表示(Connor et al,2002);总酚含量采用Folin-Ciocalteu 法测定,测定结果以没食子酸当量表示(Xu &Mou,2016);类黄酮含量采用分光光度法测定,测定结果以儿茶素水合物当量表示(Dou et al,2018);抗氧化活性采用ABTS 方法测定,测定结果以Trolox(水溶性VE)当量表示(Arnao et al,2001)。测定结果根据地上部鲜质量折算成整株含量和活性。
1.4 数据处理
试验数据采用JMP 软件整理和计算,并进行单因素方差分析和作图。
2 结果与分析
2.1 远红光对植株生长发育的影响
试验结果表明,远红光代替部分光合有效辐射使植株的株高和开展度分别提高了49%~65%和10%~17%(图1),植株地上部鲜质量和干质量分别增加了6%~23%和4%~28%(图2)。其中株高的增加值明显大于开展度,表明远红光对植株茎秆伸长的促进作用大于其对叶柄及叶片伸长的促进作用。同时,远红光处理对植株的节间数无显著影响(均在5 节左右),表明远红光主要促进了植株节间茎秆的伸长,而非节间数的增加。在红蓝光LED3 处理下,远红光处理(RBFR)提高了植株叶面积12%,但在其他光源下远红光处理对植株叶面积无显著影响(图1)。
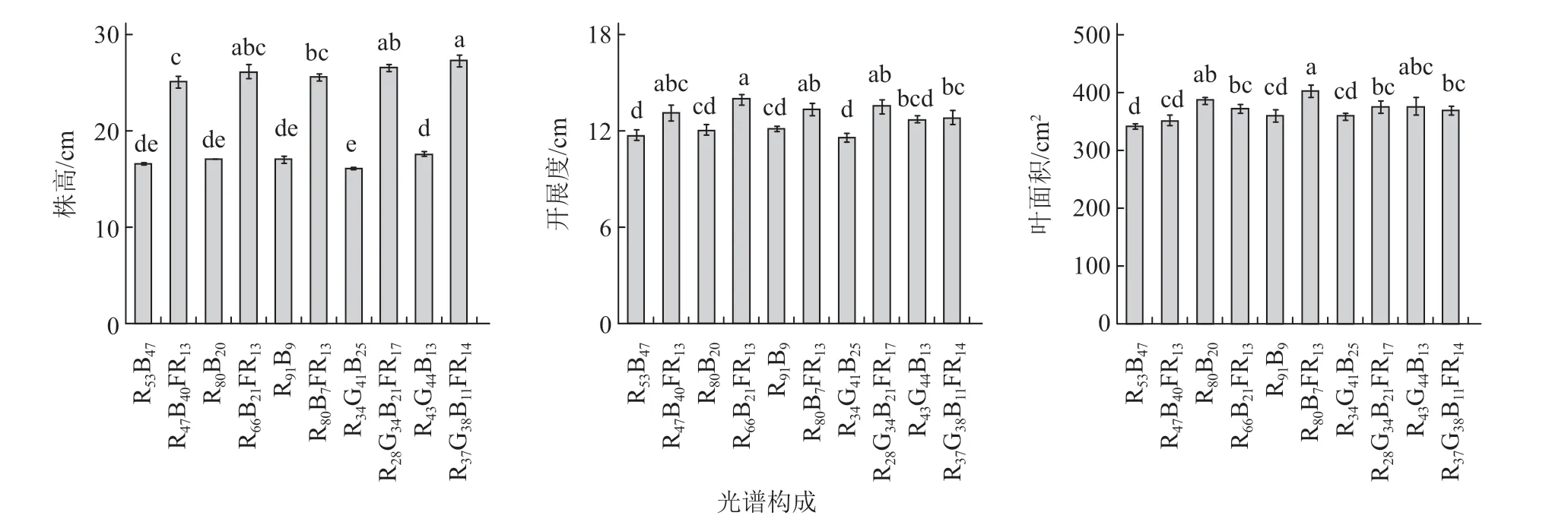
图1 不同光质处理下罗勒植株的株高、开展度和叶面积的变化

图2 不同光质处理下罗勒植株的地上部鲜质量和干质量的变化
2.2 远红光对植株光合作用和叶绿素含量的影响
在相同的总光量子通量密度下,远红光代替部分光合有效辐射对罗勒叶片的净光合速率无显著影响(图3)。保持总光量子通量密度不变,远红光代替部分光合有效辐射(RBFR、RBFR)显著降低了红蓝光LED1、红蓝光LED3 处理下罗勒叶片的叶绿素相对含量(SPAD)值,以及红蓝光LED3 处理下罗勒叶片中叶绿素a+b 的含量(图4)。

图3 不同光质处理下罗勒叶片的净光合速率的变化
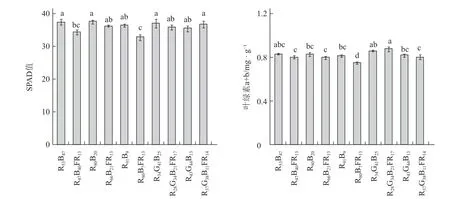
图4 不同光质处理下罗勒叶片的叶绿素相对含量和叶绿素a+b 含量的变化
2.3 远红光对植株花青素、总酚和类黄酮积累的影响
如图5 所示,在红蓝光LED2 和白色LED 处理下,远红光代替部分光合有效辐射(RBFR、RGBFR)显著提高了植株叶片花青素、总酚和类黄酮的含量。在红蓝光LED1、红蓝光LED2和白色荧光灯处理下,远红光代替部分光合有效辐射(RBFR、RBFR和RGBFR)显著提高了叶片抗氧化活性,分别提高了21%、44%和22%;且RBFR和RGBFR不同程度地提高了植株叶片花青素、总酚和类黄酮的含量。
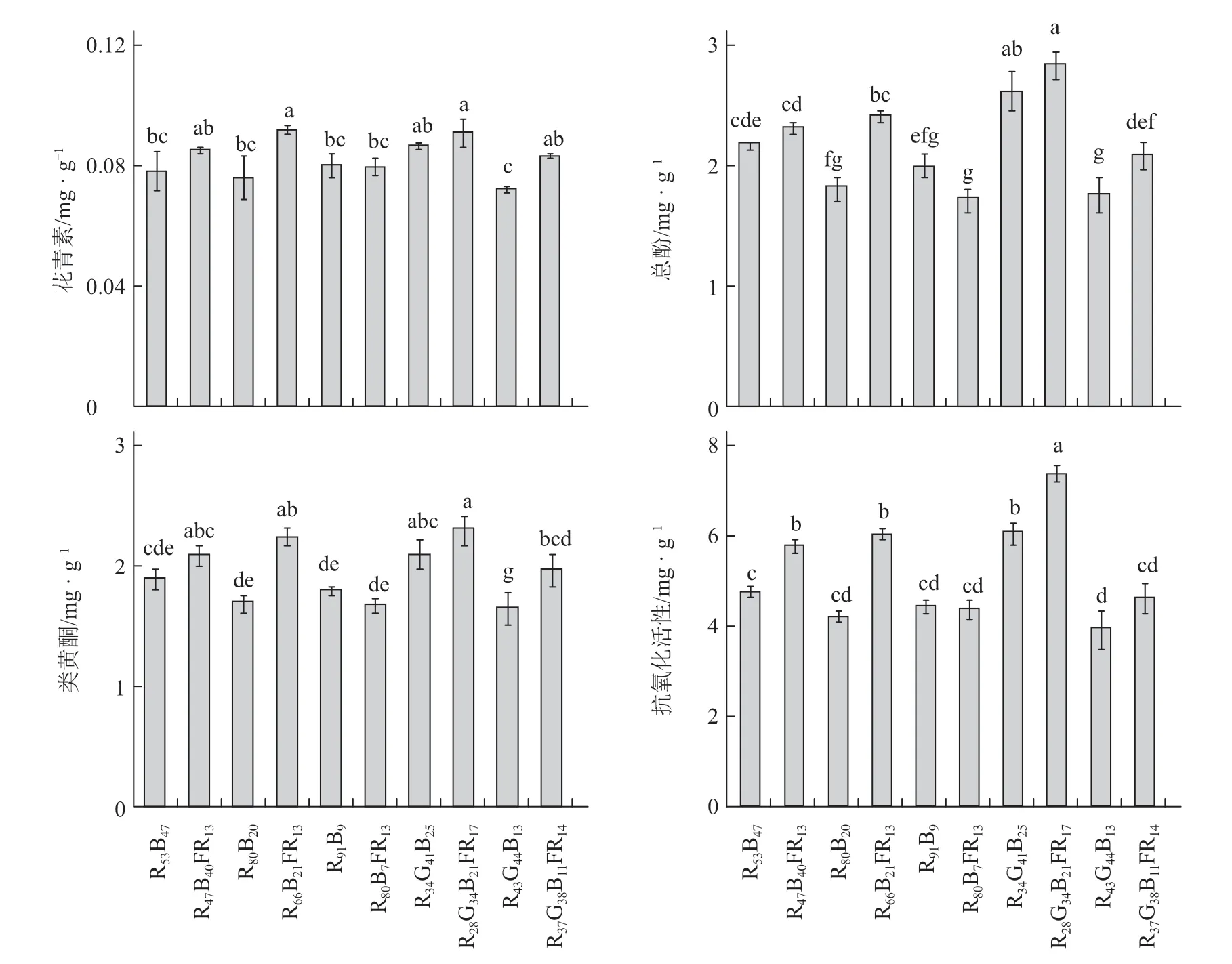
图5 不同光质处理下罗勒叶片的花青素、总酚、类黄酮含量和抗氧化活性
在红蓝光LED1 处理下,远红光代替部分光合有效辐射(RBFR)显著提高了植株花青素含量;在红蓝光LED2 处理下,远红光代替部分光合有效辐射(RBFR)显著提高了植株总酚和类黄酮含量;除红蓝光LED3 处理外,远红光代替部分光合有效辐射均显著提高了植株的抗氧化活性(表2)。

表2 不同光质处理下罗勒植株的花青素、总酚、类黄酮含量及植株抗氧化活性
3 结论与讨论
本试验中,远红光辐射下,罗勒植株的株高、地上部鲜质量和干质量提高程度高于开展度和叶面积,表明植株地上部鲜质量的增加主要是由于远红光辐射促进了植株茎秆的伸长而非叶面积的增大。研究表明,远红光辐射能够显著提高植株茎秆中赤霉素和生长素的含量,并调控植株茎秆伸长(Kurepin et al.,2007)。红蓝光LED 光源中增加远红光辐射和用远红光代替部分红蓝光均提高了天竺葵、金鱼草、叶用莴苣和羽衣甘蓝的株高和叶面积(Park &Runkle,2017),提高了叶用莴苣和羽衣甘蓝的地上部、地下部的鲜质量和干质量(Li &Kubota,2009;Meng et al.,2019)。
本试验结果表明,在相同的总光量子通量密度下,远红光代替部分光合有效辐射对罗勒植株的净光合速率无显著影响。相同地,Zhen 和Bugbee(2020)通过对12种C和2种C植物的研究表明,在相同的总光量子通量密度下,尽管植株的光合碳同化途径、冠层大小及形态、叶片结构和色素组成不同,但利用远红光代替部分光合有效辐射对植株的光合速率无显著影响。尽管植株对波长680 nm以上的辐射光量子利用效率随着波长的增加显著降低,但远红光辐射与光合有效辐射能够协同提高植株的净光合速率(Zhen &van Iersel,2017;Zhen &Bugbee,2020)。一种假设认为远红光与光合有效辐射能够协同促进PSI 和PSII 的平衡,从而弥补光合有效辐射波段损失的光量子利用效率,最终保持植株的光合效率不变或者提高植株的光合效率(Sager et al.,1989;Yang et al.,2020)。但当远红光比例高于一定值后,远红光对PSI 和PSII 的平衡无进一步的促进作用,同时由于光合有效辐射波段强度的降低对植株光合作用的抑制进一步增强,使得植株总体表现为净光合速率的降低。因此,为保证植株的光合速率,在红蓝光源中添加远红光时应将远红光的比例限定在一定范围内(Zhen &van Iersel,2017;Zhen &Bugbee,2020)。本试验中,远红光比例为13%时显著增加了植株株高,且由于株高的增加造成了部分植株倒伏。因此,在生产中决定远红光在总光量子通量密度中的比例时应综合考虑植株的净光合速率、产量、形态及植株的用途。另一种假设认为远红光辐射诱导叶绿素f进入PSI 复合物并代替部分叶绿素a,进而降低了植株叶片的叶绿素a+b 含量,为适应叶绿素f 的介入,6 个PSI 亚基被FaRLiP 基因组编码的旁系同源物所替代,进而提升了植株对远红光的利用效率(Gisriel et al.,2020)。
本试验中,远红光代替部分光合有效辐射提高了部分光源下的罗勒叶片中花青素、总酚和类黄酮的含量及叶片的抗氧化活性。Courbier 等(2020)研究表明,远红光提高了番茄叶片中有益于病原菌感染的可溶性糖含量,特别是葡萄糖和果糖,进而诱导了植株防御机制的启动,即植株次生代谢产物的积累。但有研究表明,远红光降低了植株次生代谢产物的积累(Kadomura-Ishikawa et al.,2013;Ballaré,2014;Holopainen et al.,2018)。罗勒和琉璃苣()中的迷迭香酸含量随着总光量子通量密度中远红光比例的降低而增加(Schwend et al.,2016)。红蓝光源或红蓝绿光源下的红叶生菜叶片中的花青素含量高于在红光和远红光处理下的植株(Stutte,2009)。本试验与其他研究结果的不同可能是由于不同的试验处理造成的。在远红光的试验中,植株栽培光源的光合有效辐射波段所用光源的波长构成(红光、蓝光、绿光的比例及各波段的波长范围)、远红光的波长范围、植株种类及其栽培条件(光照强度、栽培密度)均不相同,进而造成试验结果的不同。因此,远红光对植株次生代谢产物积累的影响仍有待进一步的研究。
综上所述,在相同的总光量子通量密度下,远红光代替部分光合有效辐射诱导了罗勒植株茎秆的伸长,进而提高了植株地上部的生物量积累,但对植株的净光合速率和叶面积无显著影响。同时,远红光代替部分光合有效辐射的罗勒植株的叶片和整株花青素、总酚、类黄酮含量和抗氧化活性与对照相比呈现增加的趋势。
猜你喜欢
中国药学药品知识仓库(2022年6期)2022-04-11
保健与生活(2021年13期)2021-07-14
课堂内外(小学版)(2021年5期)2021-01-17
辽河(2020年12期)2020-01-05
孩子(2019年2期)2019-02-20
中学生博览·文艺憩(2017年8期)2018-03-05
文艺生活·中旬刊(2017年10期)2017-11-19
食品与健康(2015年7期)2015-07-14
