
浙江三门湾鱼卵、仔稚鱼群聚结构时空变化研究*
2022-05-30 01:14邱科程张荣良程方平曾江宁欣冯志华陈全震徐旭丹
海洋与湖沼 2022年3期
邱科程 张荣良 晁 敏 程方平 曾江宁 彭 欣冯志华 陈全震 徐旭丹 黄 伟①
(1. 江苏海洋大学海洋科学与水产学院 江苏连云港 222005; 2. 自然资源部第二海洋研究所 自然资源部海洋生态系统动力学重点实验室 浙江杭州 310012; 3. 中国水产科学研究院东海水产研究所 上海 200090; 4. 浙江省海洋水产养殖研究所浙江温州 325005)
鱼类早期生活史阶段包括鱼卵、仔鱼和稚鱼期,是其生命中最脆弱和对栖息环境变化最敏感的阶段(Houde, 1987; 殷名称, 1991)。鱼类群聚是指生活在同一水域或生境中各种鱼类的聚合体, 鱼类早期生活史阶段的成活率直接关系到渔业资源的年际补充量, 是影响鱼类早期生活史群聚结构变化的主要因素(Chamberset al, 1997)。研究鱼卵、仔稚鱼群聚结构是评估渔业资源可持续利用度的重要组成, 对于量化海洋鱼类资源分布、有效实现鱼类物种的增殖养护、确定渔场和渔汛等都具有重要意义(Clarket al, 1953)。
海湾生境多样、饵料丰富, 有利于生物的繁殖和生长, 为海洋鱼类提供了重要产卵场、育幼场和索饵场(Archambaultet al, 1999)。鱼类早期资源的群聚组成与海湾环境有密切的联系, 良好的海湾环境有利于提高鱼类早期资源的存活率, 并在塑造鱼类群聚结构和空间分布以及物种多样性方面发挥了重要作用(Bradburyet al, 2008; Álvarezet al, 2015)。三门湾位于浙江省中部沿海, 海域面积775 km2, 是浙江省第二大海湾, 入湾河流30 多条, 其中以青溪、白溪和珠游溪最大(曾江宁等, 2011)。三门湾水质肥沃, 为鱼类产卵育肥提供了良好条件, 使其成为东海渔场的重要组成部分, 是蓝点马鲛(Scomberomorus niphonius)、鳓(Ilisha elongata)、(Liza haematocheila)等重要经济种类的产卵场, 也是浙江省鱼、虾、蟹、贝、藻等经济物种的增养殖基地, 2018 年三门湾海水养殖产量高达27.7 万t (周永东等, 2018)。20 世纪70 年代以来,随沿岸工农业发展, 高强度捕捞和环境污染导致三门湾渔业呈现资源衰退、遗传多样性水平降低的现象(宁修仁, 2005; 王星火等, 2021), 主要表现为低经济价值鱼类数量增加以及渔获个体小型化、低龄化(冀萌萌等, 2016)。此外, 一些鱼类的产卵习性也发生了变化, 表现为产卵场位移, 如小黄鱼(Larimichthys polyactis)产卵场逐渐向湾外的鱼山渔场转移(凌建忠等, 2006; 周永东等, 2011)。
目前, 关于三门湾渔业资源的研究资料十分有限, 仅开展了三门湾游泳动物种类组成、群聚结构特征及物种多样性等少量研究(蔡萌等, 2009; 李新等,2014; 冀萌萌等, 2016)。但有关三门湾鱼卵、仔稚鱼群聚结构等未见相关报道。本文基于三门湾春、夏两季调查资料, 分析了该海域鱼卵、仔稚鱼的种类组成、时空分布、群聚结构特征等, 探讨了群聚结构与环境因子的相关性。旨在了解环境变异条件下三门湾鱼类种群早期补充群体分布特征, 为科学认识渔业种群早期生活史补充过程提供基础资料和数据支撑。
1 材料与方法
1.1 调查站位和方法
三门湾三面环陆, 是呈西北-东南走向的半封闭海湾, 在该海域设置20 个调查站位(121.45°~122.18°E,28.65°~29.36°N), 根据底质类型、水文条件将其分为湾内(1~10)、湾外(11~20)两个区域(图1) (聂振林等,2018)。分别开展夏(2019 年8 月)、春(2020 年4 月)两季的鱼卵、仔稚鱼资源调查。依照《海洋调查规范》(中华人民共和国国家质量监督检验检疫总局等,2018), 采用浅水Ⅰ型浮游生物网(网口面积0.2 m2,网口直径50 cm, 网长145 cm)定量采样, 由海底至海面进行垂直拖网, 网口均配备流量计(Hydro-Bios-438115, 德国)记录拖网流量值; 采用大型浮游生物网(网口面积0.5 m2, 网口直径80 cm, 网长280 cm)定性采样, 水平拖网10 min, 平均拖速2 kn。同时, 使用多参数水质分析仪(YSI-Pro, 美国)现场测量水温、pH、溶解氧、盐度等环境参数。样品用5%中性甲醛溶液固定保存, 待返回实验室后进行挑选、分类鉴定并计数, 物种鉴定主要参考《中国近海鱼卵与仔鱼》(张仁斋, 1985)、《中国近海及其临近海域鱼卵与仔稚鱼》(万瑞景等, 2016)、《长江口鱼类》(庄平等, 2006)等资料。鱼卵、仔稚鱼的数量分布以每一站位实际采获的个体数表示, 单位分别为粒和尾。

图1 三门湾鱼卵、仔稚鱼调查站位Fig.1 Sampling stations of fish eggs, larvae and juveniles in the Sanmen Bay
1.2 数据统计分析
(1) 鱼卵、仔稚鱼密度G(ind./m3)

式中,G为单位体积海水中鱼卵(粒/m3)或仔稚鱼个体数(尾/m3);Na为全网鱼卵或仔稚鱼个体数;S为网口面积(m2);L为流量计转数;C为流量计校正值(C=0.3)(国家海洋局908 专项办公室, 2006)。
(2) 物种优势度及更替率
用优势度(Y)确定鱼卵、仔稚鱼的优势种类, 其计算公式为:

式中,ni为第i种鱼的鱼卵(或仔稚鱼)数量,N为所有种类的鱼卵(或仔稚鱼)总数量,f为站位出现频率, 取优势度大于0.02 的为优势种(McNaughtonet al, 1967)。
使用种类组成更替率(R)来衡量鱼卵、仔稚鱼种类组成的季节更替情况, 其公式为:

式中,a和b分别表示相邻两个季节的物种的种类数,c为两个季节共有物种的种类数(刘瑞玉, 1992)。
(3) 群聚结构分析
采用Primer 6.0 软件进行聚类分析, 为平衡优势种和非优势种在群聚中的作用, 计算前对原始数据矩阵进行lg(x+1)变换, 再计算Bray-Curtis 距离, 建立距离矩阵, 最后采用类平均法进行聚类并作树状图(Clarke, 1993; Brazneret al, 1997)。为了验证聚类结果的可信度, 在此基础上进行了非线性多维标度排序分析(Nonmetric Multidimensional Scaling, NMDS);应用相似性检验(Analysis of similarities, ANOSIM)非参数多元分析方法(Clarke, 1993)检验群落之间的差异显著性; 应用相似性百分比法(Similarity Percentage,SIMPER)分析鱼类对站位组内相似性和组间相异性的贡献率; 用胁强系数(stress)来衡量NMDS 结果的优劣, 此值越小越好, 当stress<0.2 时, 可用NMDS的二维点图表示, 其图形有一定的解释意义; 当stress<0.1 时, 可以认为是一个好的排序; 当stress<0.05时, 具有很好的代表性(Khalafet al, 2002)。
(4) 环境因子分析
采用Canoco 5.0 分析鱼类物种与环境因子之间的关系并绘制排序图, 以揭示不同环境因子对鱼卵 、仔稚鱼的影响。首先对物种数据进行消除趋势对应分析(Decentred Correspondence Analysis, DCA), 根据每个轴的梯度长度(Length of Gradient Axis, LGA)选择适宜的排序方法。当 LGA<3 时, 选择冗余分析(Redundancy Analysis, RDA); 当LGA>4 时, 则可运用典范对应分析(Canonical Correspondence Analysis,CCA); 当3<LGA<4 时, 两者皆可(Lepšet al, 2003)。根据DCA 分析结果, 本文采用线性模型的RDA 分析。
采用 SPSS 23.0 软件进行单因素方差分析(One-way ANOVA), 检验物种丰度时空差异性。
2 结果
2.1 种类及优势种组成
两季共鉴定出鱼卵、仔稚鱼8 目21 科40 种, 其中定性样品鉴定出鱼卵、仔稚鱼8 目21 科36 种, 定量样品共4 目11 科24 种(详见附录)。定量样品中鲈形目(Perciformes)种类最多, 鉴定出7 科17 种, 占总采获量的51.9%; 鲽形目(Pleuronectiformes)最少, 占总采获量的10.6%。春季调查中, 鉴定出鱼卵4 种, 隶属于3 目4 科, 仔稚鱼6 种, 隶属于3 目5 科, 鱼卵、仔稚鱼共有种3 种; 夏季调查中, 鉴定出鱼卵5 种,隶属于3 目5 科, 仔稚鱼13 种, 隶属于3 目6 科, 鱼卵、仔稚鱼共有种1 种; 夏季鱼卵、仔稚鱼采获量均最多, 分别占总采获量的20.6%和51.3%; 鱼卵采获量最少的是春季, 占总采获量的7.5%。

附录 三门湾春、夏季鱼卵、仔稚鱼种类组成Appendix Species composition of fish eggs, larvae and juveniles in spring (2020) and summer (2019) in the Sanmen Bay

续表
根据优势度(Y)大于0.02 为优势种的原则, 该调查海域鱼卵和仔稚鱼优势种共有10 种, 优势度最高的物种为(见表1)。其中春季鱼卵样品中优势种为斑(Konosirus punctatus)、鲷科未定种(Sparidae)、蓝点马鲛、, 各占春季鱼卵样品的25% (见图2); 仔稚鱼样品中优势种为, 占春季仔稚鱼样品的76%。夏季鱼卵样品中优势种为舌鳎属(Cynoglossusspp.)、石首鱼科未定种(Sciaenidae)、小公鱼属(Stolephorusspp.), 分别占夏季鱼卵样品的39%、27%、21%; 仔稚鱼样品中优势种为孔虾虎鱼、小公鱼属、细棘虾虎鱼属(Acentrogobiusspp.)、缟虾虎鱼属(Tridentigerspp.),分别占夏季仔稚鱼样品的35%、18%、12%、11%。从生态类型上看, 春季鱼卵、仔稚鱼优势种主要是河口性鱼类; 夏季鱼卵优势种主要是海洋性鱼类, 仔稚鱼优势种主要是河口性鱼类。从空间上看, 湾内春、夏两季优势种共有10 种, 湾外为5 种; 湾内鱼卵、仔稚鱼优势种主要为河口性鱼类; 湾内、湾外共有优势种4 种(见表2)。
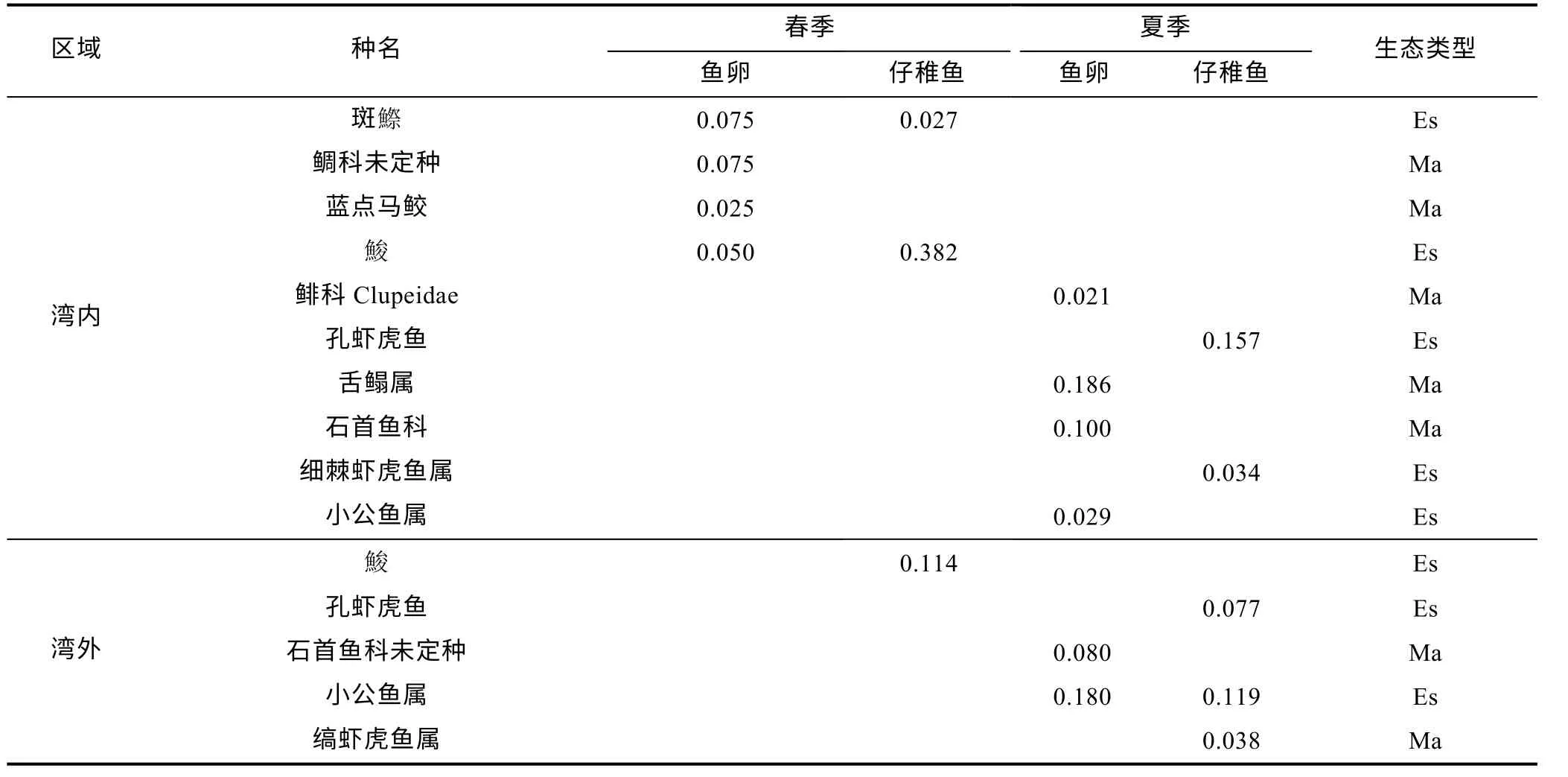
表2 三门湾湾内、湾外鱼卵、仔稚鱼优势种组成Tab.2 Dominant species composition of fish eggs, larvae and juveniles inside and outside the Sanmen Bay

图2 三门湾鱼卵、仔稚鱼优势种季节占比Fig.2 Seasonal proportion of dominant species of fish eggs, larvae and juveniles in the Sanmen Bay

表1 三门湾春、夏两季鱼卵、仔稚鱼优势种组成及生态类型Tab.1 Dominant species composition and ecological types of fish eggs, larvae and juveniles in the Sanmen Bay in spring and summer
2.2 密度时空变化
三门湾鱼卵、仔稚鱼密度的季节及空间变化如图3 所示, 统计结果显示, 春季鱼卵、仔稚鱼的平均密度均显著低于夏季(ANOVA,P<0.05), 其中春季鱼卵和仔稚鱼的平均密度分别为0.40 ind./m3和1.04 ind./m3;夏季鱼卵和仔稚鱼的平均密度分别为0.62 ind./m3和1.70 ind./m3; 此外, 图3a 中出现的异常值, 最大值为夏季仔稚鱼5 号站位。空间分布见图3b, 湾内春、夏两季鱼卵、仔稚鱼密度均高于湾外(1.34 ind./m3vs0.54 ind./m3), 除了夏季湾内仔稚鱼密度与湾外比较无显著差异(P>0.05)外, 春季湾内鱼卵、仔稚鱼密度及夏季湾内鱼卵密度与湾外比较均具有显著差异(P<0.05)。

图3 三门湾鱼卵、仔稚鱼密度季节及空间变化Fig.3 Seasonal and spatial variation in the density of fish eggs, larvae and juveniles in the Sanmen Bay
2.3 群聚结构分析
三门湾20 个调查站位鱼卵、仔稚鱼群聚结构季节变化聚类分析结果如图4 所示(其中春季5、6、8、13~18 号站, 夏季2、14、18、20 号站均无采获, 为减少误差故不作统计)。春、夏两个季节的群聚划分明显(7.4%的相似性水平), 相似性检验(ANOSIM)非参数多元分析结果显示2 个群聚差异极显著(R=0.589,P<0.01), 反映出鱼卵、仔稚鱼群聚结构的季节演替明显(图4a)。NMDS 排序的胁强系数为0.01 (stress<0.05),说明NMDS 排序图具有很好的代表性(图4b)。两季物种更替率R达到100%, 进一步说明了三门湾鱼卵、仔稚鱼群聚结构季节变化具有极显著的差异。另外,结合相似性百分比(SIMPER)分析, 结果显示春季形成的湾内、湾外群聚结构差异显著(P<0.05) (图4c),主要分歧种有、斑、矛尾虾虎鱼(Chaemrichthys stigmatias)等, 其组间相异性贡献率达到76.5% (表3); 而夏季湾内、湾外群聚结构无显著差异(P>0.05)(图4d)。

表3 三门湾湾内、湾外鱼卵、仔稚鱼分歧种组成Tab.3 Composition of of fish eggs, larvae and juveniles of different species inside and outside the Sanmen Bay

图4 三门湾鱼卵、仔稚鱼群聚结构季节变化聚类图、排序图Fig.4 Clustering and sorting charts of seasonal changes in cluster structures of fish eggs, larvae and juveniles in the Sanmen Bay
2.4 环境因子分析
三门湾海域鱼卵、仔稚鱼优势种与环境因子的RDA 分析结果如图5 所示。春季, 5 个环境因子解释了物种-环境关系的45.8%, 第一RDA 轴(RDA1, 特征值=0.425)和第二RDA 轴(RDA2, 特征值=0.012)表现出很高的物种-环境相关性(RDA1 和RDA2 与环境因子相关性分别为0.705 和0.458)。蒙特卡洛显著性检验结果显示, 第一排序轴和所有排序轴均差异不显著(第一轴:F=10.3,P=0.09; 所有轴:F=2.2,P=0.09)。影响三门湾春季鱼卵、仔稚鱼群聚的主要环境因子是水温、pH、溶解氧, 其中第一排序轴与水温呈最强正相关, 与水深呈负相关, 第二排序轴与溶解氧呈正相关, 与水深呈最大负相关。结果还表明斑鲫、等鱼卵、仔稚鱼受水温影响较大, 鲷科未定种鱼卵、仔稚鱼受pH 影响较大。夏季, 5 个环境因子解释了物种-环境关系的52.3%, 第一RDA 轴(RDA1,特征值= 0.187)和第二RDA 轴(RDA2, 特征值=0.038)表现出良好的物种-环境相关性(RDA1 和RDA2 与环境因子相关性分别为0.569 和0.523)。蒙特卡洛显著性检验结果显示, 第一排序轴和所有排序轴差异均不显著(第一轴:F=3.2,P=0.39; 所有轴:F=1.0,P=0.37)。影响三门湾夏季鱼卵、仔稚鱼群聚的主要环境因子是pH、水温、盐度, 其中第一排序轴与pH呈最强正相关, 与水温呈负相关, 第二排序轴与盐度呈正相关, 与水深呈最大负相关。从物种和环境因子关系来看, 孔虾虎鱼鱼卵、仔稚鱼受pH 响较大, 舌鳎属、缟虾虎鱼属受温度影响较大。

图5 三门湾海域鱼卵、仔稚鱼优势种密度与环境因子之间的关系Fig.5 Relationship between density of dominant species of fish eggs, larvae and juveniles and environmental factors in the Sanmen Bay
3 讨论
3.1 鱼卵、仔稚鱼组成及优势种
本研究中定量样品鉴定出的鱼卵、仔稚鱼隶属于4 目11 科24 种, 其中鲈形目种类最多, 为7 科17 种,占总采获量的51.9%, 与舟山渔场(鲈形目共19 种,占47.50%)海域研究结果相似(刘连为等, 2021)。三门湾鱼卵主要优势种为斑、小公鱼属、石首鱼科未定种等, 仔稚鱼主要优势种为、小公鱼属等, 与象山港、岱衢洋和瓯江口等邻近海域的类似(杜萍等, 2016;王宇坛等, 2017; 樊紫薇等, 2020)。但与长江口冲淡水区域群落差别较大, 后者以 鳀、凤鲚等为主要优势种(表4) (吴光宗, 1989; 杨东莱等, 1990)。
20 世纪50~60 年代, 三门湾渔业种类主要是小黄鱼、带鱼(Trichiurus lepturus)和鲆鲽类等经济价值较高的底层、近底层鱼类(Jin, 2004)。20 世纪70~80年代以后, 违反自然规律的过度捕捞已远超渔业资源的再生能力, 使经济价值较高的鱼类资源量不断下降(凌建忠等, 2006)。此外, 围填海、海底管线等近岸工程, 进一步破坏了三门湾传统渔业生物的产卵场和栖息地, 也会影响渔业资源补充过程(黄潘阳等,2017)。本调查采集到的鱼卵、仔稚鱼以小型低值鱼类如虾虎鱼、斑、小公鱼属、鲱科未定种、舌鳎属、石首鱼科为主, 带鱼、鲆鲽类等经济价值较高、营养级较高的种类未出现或数量较少, 表明该海域优势种可能存在由多种共存逐渐向单种为主转变, 由经济价值较高的鱼类向低经济价值以及小型化鱼类转化的趋势(张波等, 2004)。
本调查中鱼卵、仔稚鱼密度与长江口、舟山渔场、岱衢洋等邻近海域相近(林楠等, 2016; 樊紫薇等,2020; 刘连为等, 2021)。尽管从“代际”中时间尺度看, 三门湾渔业资源严重衰退, 但从“年际”短时间尺度上看, 本次调查仔稚鱼密度较2013 年(仔稚鱼0.49 ind./m3)已有回升趋势(表4)。其原因可能是: 近年来三门湾周边市县实施限制捕捞规格、伏季休渔期、增殖放流等相关政策, 尤其是2017 年史上最严休渔制度的实施发挥了积极作用, 仔稚鱼密度变化趋势反映了三门湾渔业资源正逐步恢复(邵晨等,2018)。三门湾增殖放流时间较早, 从20 世纪90 年代即开始放流对虾, 至2019 年, 三门湾周边市县增殖放流苗种达2.4 亿尾(头)/年, 主要品种为大黄鱼、黑鲷、黄姑鱼、斑等(朱维新, 1990; 周永东, 2004)。另一方面, 从三门湾游泳动物生物量资源密度变化来看, 2008 年(175.32 kg/km2)、2013 年(208.37~213.34 kg/km2, 中国水产科学研究院东海水产研究所,2014)、2015 年(314.44 kg/km2)、2020 年(356.73~384.10 kg/km2), 已初步呈逐年恢复趋势(蔡萌等,2009; 冀萌萌, 2016; 马路阔等, 2022)。

表4 三门湾海域及临近海域鱼卵、仔稚鱼密度及优势种的比较Tab.4 The abundance comparison of fish eggs, larvae and juveniles in in the Sanmen Bay and adjacent area
3.2 鱼卵、仔稚鱼群聚的空间异质性
本次调查湾内采获的鱼类优势种多于湾外, 湾内采获的主要为鱼卵。从空间分布上看, 湾内外春、夏两季鱼卵、仔稚鱼的群聚结构差异显著, 且湾内的资源密度均高于湾外。导致这一空间分布格局的主要原因为: 首先, 鱼类优势种的生活习性, 尤其是产卵时的生境偏好导致湾内外的资源量差异。SIMPER 分析结果显示,、斑、矛尾虾虎鱼等优势种是导致湾内外鱼卵、仔稚鱼群聚结构差异的主要分歧种(表3)。这些种类主要是河口性鱼类, 营养级较低, 产卵量大, 温盐度适应性强, 常年栖息于湾内, 同时鱼卵发育成仔鱼的周期较短(大多数仅需1~3 d, 稍长的最多5 d), 在湾内便发育成仔鱼, 随海流扩散的现象不明显, 这一结果也印证了优势种对于群聚结构的塑造作用。其次, 三门湾是一个半封闭性港湾, 受外海海流影响较小, 同时涉海工程如围涂进一步缩减了湾内滩涂面积, 使湾内纳潮量减小, 导致流速亦相应减小, 水体滞留时间增长(黄潘阳等, 2017)。由于鱼卵不具备活动能力, 随着波浪漂流, 故湾内鱼卵随海流扩散的现象不明显(王建茹等, 2010)。最后, 三门湾流域咸淡水交汇充分的湾内水域, 大量陆源输入的营养盐会刺激藻类的生长, 进而通过食物链间接为鱼类提供了丰富的饵料, 因而相比湾外更可能成为鱼类理想的产卵场(曾江宁等, 2011)。
另一方面, 鱼卵和仔稚鱼的生长发育、数量分布等与海流、盐度以及水温等环境因子密切相关(Milleret al, 2009)。环境因子分析结果表明本研究中引起三门湾海域鱼卵、仔稚鱼群聚结构空间变化的主要因子是温度。其中, 春季湾内平均水温(18.44 °C)明显高于湾外(16.95 °C), 根据Hurst 等(2010)对鱼卵生长动力学研究, 鱼类在产卵期和仔鱼期的大小与温度呈正相关。春季鱼卵、仔稚鱼优势种斑、蓝点马鲛、等种类所占比例较大, 且多为暖温(水)性中上层鱼类,对环境变化的敏感度较高, 受水温影响较大, 由此形成了湾内物种多样且产卵频繁的空间特征。夏季, 尽管湾内平均水温(29.50 °C)略低于湾外(29.70 °C), 但舌鳎属、缟虾虎鱼属、孔虾虎鱼等都为底层鱼类, 此外, 湾内饵料丰富, 鱼卵发育成仔、稚鱼后可就地索饵。因此, 形成了湾内鱼卵、仔稚鱼密度高于湾外的分布格局。本研究中, 斑、鲷科未定种、等分布还与pH 呈正相关, pH 主要通过影响仔稚鱼的嗅觉等调控鱼类生长发育(Mundayet al, 2009)。研究表明pH≥8.1 时, pH 与鱼卵、仔稚鱼丰度呈负相关(王九江等, 2019)。本研究中湾内pH (8.07)略小于湾外(8.15),更适宜仔稚鱼生活。盐度对鱼卵、仔稚鱼分布的影响与鱼类本身特性有关, 鱼卵、仔稚鱼对盐度的适应范围较窄, 对盐度的适应性随着仔稚鱼的生长逐渐增强(黄永春等, 2006)。本研究中盐度对鱼卵、仔稚鱼的影响并不明显, 这可能是由于本调查采集的主要种类斑、鲷科未定种、等是广盐性鱼类, 对盐度有较强的耐受性。
3.3 鱼卵、仔稚鱼群聚的季节变化
群聚结构季节变化聚类分析显示, 调查海域鱼卵、仔稚鱼组成存在明显的季节变化, 与长江口(蒋玫等, 2014)、杭州湾(闫欣等, 2014)、舟山群岛外海域(于南京等, 2020)等临近海域鱼卵、仔稚鱼群聚结构季节变化结果相符(陈渊戈等, 2015; 樊紫薇等, 2020)。一方面, 鱼类产卵存在特定的时间节律。通常鱼类的产卵期为该海域鱼卵大量集中出现的时期(赵优等,2008), 夏季三门湾的鱼卵、仔稚鱼丰度均达到最高值,意味着该季节多数鱼类进入产卵期。春、夏两季节鱼卵、仔稚鱼物种(尤其是优势种)更替明显, 且三门湾往年渔业资源调查结果显示, 成鱼的优势种更替情况与之相符(冀萌萌等, 2016; 聂振林等, 2018; 俞松立等, 2021)。另一方面, 长距离洄游物种(如蓝点马鲛), 清明节前后进入湾内产卵并滞留到秋季, 其稚鱼主要在岛屿及岩礁附近海域索饵, 且夏季通常栖息于中上层, 故容易被采获。舌鳎属每年12 月起向湾外东部海域越冬洄游, 翌年2 月向浙江沿海产卵洄游, 主要产卵期是5~8 月, 且主要产卵场在湾外(周永东等, 2018)。这些鱼类的产卵洄游习性很大程度上影响了群聚的季节变化。
此外, 三门湾属暖温带海域, 受陆地径流、江浙沿岸流及台湾暖流等水团共同影响, 季节差异明显,如海域潮流场流速的季节变化为夏季略大于冬季,秋季略大于春季(曾江宁等, 2011); 寒暖流交汇使海水扰动, 下层营养物质带到表层, 有利于鱼类繁殖索饵, 相互作用下形成鱼卵、仔稚鱼群聚的季节差异(张启龙等, 2007)。
4 结论
(1) 三门湾海域鱼卵、仔稚鱼的数量及分布现状:两季共采获到鱼卵、仔稚鱼8 目21 科40 种, 其中定性样品共采获鱼卵、仔稚鱼8 目21 科36 种, 定量样品共采获鱼卵、仔稚鱼4 目11 科24 种。总体而言, 三门湾鱼卵、仔稚鱼资源密度与周围海域相近, 优势种种类差异不大。
(2) 空间分布上, 三门湾湾内与湾外鱼卵、仔稚鱼群聚的组成存在明显差异。主要由鱼类优势种的生活习性以及产卵时的生境偏好导致湾内外的资源量差异。SIMPER 分析结果显示, 其群聚空间差异主要由、斑、矛尾虾虎鱼等分歧种引起。此外, RDA分析结果显示, 引起三门湾海域鱼卵、仔稚鱼群聚结构空间变化的主要环境因子是温度。
(3) 季节变化上, 三门湾春夏两季鱼卵、仔稚鱼优势种更替明显。春季鱼卵、仔稚鱼优势种主要为斑、鲷科未定种、蓝点马鲛、; 夏季鱼卵、仔稚鱼优势种主要为舌鳎属、石首鱼科未定种、小公鱼属等。
猜你喜欢
水产科学(2022年1期)2022-01-26
中国宝玉石(2021年5期)2021-11-18
浙江海洋大学学报(自然科学版)(2021年1期)2021-07-06
儿童时代·幸福宝宝(2020年9期)2020-09-08
创新作文(3-4年级)(2019年8期)2019-12-17
孩子·小学版(2019年2期)2019-09-10
兵器知识(2017年10期)2017-10-19
探索科学(2017年5期)2017-06-21
环球时报(2016-03-22)2016-03-22
人民周刊(2016年1期)2016-01-09
