
干旱胁迫及复水对“雪不凋”萱草生理特性的影响研究
2022-06-02 03:29王晓红黄程前郑硕理李立华陈佳佳胡灵璇
绿色科技 2022年9期
方 娟,王晓红,黄程前,郑硕理,李立华,陈佳佳,胡灵璇
(1.中南林业科技大学,湖南 长沙 410004;2.湖南省森林植物园,湖南 长沙 410004)
1 引言
随着全球气候变化引起的温度升高、降雨增多和干旱频发,且干旱区“干湿交替”现象加剧,非干旱地区也存在季节性干旱现象[1]。湖南省内各地区季节性干旱严重,且降雨量分布不均匀,尤其是夏秋高温少雨天气导致的气象干旱会限制植物生长发育。干旱胁迫会引起细胞失水,破坏植物体内水分代谢,导致植物体生理生化和光合特征发生重大变化[2],甚至导致植物死亡。当植物经过一定程度的干旱胁迫后,对其进行复水,一般认为植物会利用其体内的调节机制进行自我修复[3]。因此研究土壤水分亏缺和旱后复水条件下对植物的生理生化及光合生理等方面的影响,探索植物体内的相关响应机制,不仅能揭示植物对干旱的适应能力和抗旱机理,同时对植物的繁殖栽培及应用有着重要意义。
萱草属植物(Hemerocallisspp.)是多年生宿根花卉,全属植物14种,我国11种和部分自然杂交变种[4,5]。萱草属植物性强健、耐寒、耐热、耐盐碱、耐半阴、适应力强、花色鲜艳、花期长,对土壤要求不严,园林应用中多用于丛植、路旁栽植或做地被应用[6]。贾民隆[7]等采用田间自然干旱的方式对萱草进行干旱胁迫;欧敏哲等[8]对大花萱草进行持续干旱及复水的生理情况,筛选出两个自我修复能力较强的大花萱草;李昊等[9]建立耐干旱萱草品种筛选的简易方法。但是对于旱后复水条件下萱草的生理生化及光合作用响应机制的研究鲜有报道,本试验中选择观赏价值较高、湖南生长良好、抗逆性普遍较强的“雪不凋”萱草进行试验,揭示“雪不凋”萱草的耐旱性的生理及光合机制。可以为“雪不凋”萱草品种的园林推广及其在干旱、半干旱地区的应用上提供参考。
2 材料与方法
2.1 实验材料
供试材料为“雪不凋”萱草取自湖南省森林植物园萱草资源圃的多年生分株苗。
2.2 实验方法
2020年5月份将萱草统一移植到上口径18 cm、下口径14.5 cm、高18.5 cm的塑料花盆中,栽培基质为珍珠岩与泥炭土(1∶1)的混合基质,每盆干土质量为(1.6±0.2)kg,每盆种植一株。期间进行正常水肥管理,促其成活。
试验采取温室盆栽控水法,选取长势整齐且健康的试验材料各45盆,随机分成5组,每组9盆。其中对照(CK):每天浇水浇透,相对含水量为(65%±2%);轻度干旱胁迫(LD):相对含水量为(50%±2%);中度干旱胁迫(MD)相对含水量为(35%±2%);重度干旱胁迫(SD)相对含水量为(20%±2%);自然干旱胁迫(ND):试验开始时浇透水后不再浇水,直至干旱结束。控水期间,CK和ND进行正常水分管理,其他处理自然干旱达到预设梯度后维持其含水量。试验期间每天17:00用电子秤称重和高精度土壤水分测试仪JK-200F测量两种方法对试验材料控水,使含水量维持在各预定胁迫水平。干旱胁迫试验持续21 d后开始复水。于旱胁迫 0、7、14、21 d和复水后第7 d采集叶片样品,用于测量指标。
2.3 指标测定
相对电导率采用电导仪[10]测定;叶绿素含量测定采用丙酮提取法[11];MDA含量采用硫代巴比妥酸法测定;超氧化物歧化酶(SOD)活性的测定采用氮蓝四唑(NBT)光化还原反应[12];游离脯氨酸的测定采用磺基水杨酸法;可溶性蛋白含量采用考马斯亮蓝法测定[3]。
光响应曲线采用便携式光合系统测定仪LI-6400 ( LI-COR Lincoln,USA)于 干旱21 d和复水7 d时的9:00~17:00,选取选择受光方向一致的萱草成熟叶片在光照强度1000 μmol/(m2·s)条件下测定光合参数,测定时由红蓝光源提供不同的 PAR:2000、1500、1200、1000、800、600、400、200、1000、50、20、0 μmol/(m2·s)。测定前对每个叶片进行20 min的光诱导, 叶室CO2浓度为380 μmol/mol。
2.4 数据处理
通过修正直角双曲线模型对萱草叶片光合作用-光响应曲线进行模拟。基本数据处理采用Excel进行计算与统计,光响应曲线采用光合计算进行双曲线修正模型模拟;用SPSS26.0进行数据分析,异性显著水平为P<0.05用Origin2018制作图表。
3 结果
3.1 干旱胁迫和复水对“雪不凋”萱草生理指标的影响
3.1.1 干旱胁迫和复水对“雪不凋”萱草相对电导率、丙二醛含量的影响
由图1a和图1b可知,“雪不凋”萱草叶片相对电导率、丙二醛含量随着胁迫程度的加剧和胁迫时间的持续而不断上升。胁迫21 d后,各处理组相对电导率都明显上升,LD、MD、SD和ND分别比CK高21.89%、66.52%、 95.43% 和106.56%且均与CK差异显著(P<0.05);其中,LD和MD的丙二醛含量缓慢升高,SD和ND则表现为迅速升高,胁迫21 d时,LD、MD、SD和ND的丙二醛含量分别比CK高38.23%、64.26%、81.36%和91.62%且均与CK差异显著(P<0.05)。
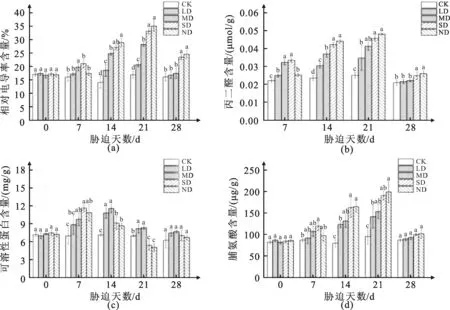
图1 干旱和旱后复水对“雪不凋”萱草相对电导率、丙二醛、可溶性蛋白、脯氨酸含量的变化
复水7d后,相对电导率和丙二醛含量和迅速降低,LD和 MD基本上恢复到对照水平,无明显差异(P>0.05),但SD和ND仍高于CK,且有显著差异,而各胁迫水平丙二醛含量基本上恢复到对照水平,与CK的差异均不显著。
3.1.2 干旱胁迫和旱后复水对“雪不凋”萱草可溶性蛋白含量、脯氨酸含量的影响
由图1可以看出,在试验时间段内,随着干旱胁迫时间的增加,各处理组均表现出先上升后下降的趋势,而各胁迫度的脯氨酸含量则表现为持续升高趋势。在干旱处理21 d 时,LD和 MD处理下的萱草叶片的可溶性蛋白含量均显著高于CK,分别比CK高16.80%、19.25%;SD和ND处理的萱草叶片的可溶性蛋白则显著低于CK,与CK相比,分别下降了22.62%、27.51%。干旱0~7 d时,LD和ND脯氨酸含量与CK差异不显著,当干旱21 d时;各处理组的脯氨酸含量均上升为最大值且显著高于CK,分别比CK增加了48.40%、60.99%、99.71%、108.42%;干旱21 d,萱草叶片脯氨酸含量下降,但仍显著高于CK。复水7 d后,萱草可溶性蛋白及脯氨酸含量的LD和MD均恢复到对照水平。
3.1.3 干旱和旱后复水对“雪不凋”萱草SOD活性的影响
如图2a可知,在干旱胁迫期间,CK的SOD活性基本保持稳定,萱草干旱SOD活性整体呈先升高后降的上升趋势。在干旱胁迫第14d时,LD和MD的SOD活性达到最大,分别比CK显著高出47.21% 、69.52% 。在胁迫21 d后,各处理组的SOD活性均显著低于对照水平。复水7 d后,各胁迫水平SOD活性基本上恢复到对照水平,与CK的差异均不显著。

图2 干旱和旱后复水对“雪不凋”萱草SOD活性、叶绿素、光响应曲线的变化
3.2 干旱胁迫和旱后复水对“雪不凋”萱草光合作用的影响
3.2.1 干旱胁迫和旱后复水对“雪不凋”萱草叶绿素含量的影响
由图2b可知,“雪不凋”萱草叶片中叶绿素含量随着干旱胁迫程度的加剧和胁迫时间的延长呈现先上升后下降的趋势或者下降趋势。其中LD和MD萱草的叶绿素含量是在干旱胁迫14 d后达到最大值,分别CK的116.15%、111.95%。在胁迫21 d后,各胁迫处理的叶绿素含量均有所下降,其中SD和ND处理组叶绿素含量分别为CK的138.99%、145.35%,且各干旱处理都和CK差异显著(P<0.05)。复水7 d后,叶绿素含量迅速上升,LD和MD基本上恢复到对照水平,无明显差异(P>0.05),但SD和ND仍高于CK,且有显著差异。
3.2.2 干旱胁迫和旱后复水对“雪不凋”萱草光响应曲线的影响
如图2c和2d所示,无论是干旱胁迫21 d,还是复水7 d后,萱草各处理组的Pn随着光强的增加逐渐升高,变化趋势均表现为低PAR(0~600 μmol/(m2·s))时Pn迅速增加,达到一定的PAR(600~1200 μmol/(m2·s))之后,Pn增幅变小,之后逐渐稳定并出现光抑制现象随着干旱程度的加剧,曲线变化幅度表现为CK>LD>MD>SD>ND。不同的是,干旱21 d后,各处理与CK差异明显,复水后,各组处理的变化曲线慢慢增大,与CK差异越来越小。
4 讨论
干旱胁迫下,植物体内的活性氧积累会对细胞膜系统造成伤害,相对电导率可以用来判断细胞膜受到损伤的严重程度[13]。丙二醛的积累和细胞膜透性变化可以反映细胞膜脂过氧化作用强弱和质膜破坏强弱[14]。试验发现,干旱胁迫下,萱草丙二醛含量和相对电导率显著上升,说明萱草叶片的膜脂过氧化程度加深,对细胞膜结构与功能造成了严重伤害,且叶片的丙二醛含量和相对电导率成正相关,这与何凤[15]等对于杜仲(EucommiaulmoidesOliv.)扦插苗的研究结果相一致。复水后,轻度和重度干旱均能恢复到正常水平,而重度干旱还是显著高于对照,这说明重度胁迫下,萱草叶片膜系统受到的伤害较大,短时间内难以恢复到正常水平。
渗透调节物质的积累是植物适应干旱胁迫的基本特征之一,其含量的积累可以在一定程度上表明植物渗透能力的强弱[16]。试验中,萱草叶片重度胁迫的可溶性蛋白和脯氨酸随着干旱时间的延长先增高后下降,这说明干旱前期,萱草主动积累可溶性蛋白和脯氨酸含量发来降低渗透压,增加了生物大分子的亲水性[17],以此抵抗干旱胁迫带来的伤害。在试验后期,可能是持续胁迫下对萱草产生了一定的伤害,使萱草达到了耐受阈值,抑制了其蛋白质和脯氨酸的合成,这与靳月等[18]闽楠(Phoebebournei)的研究结果基本一致。在复水7 d后,各干旱处理水平均恢复为对照水平,说明萱草具有较强的耐旱性。
SOD被认为是ROS清除系统中的关键酶,是清除ROS的第一道防线[19],其活性变化与膜脂过氧化程度有密切联对活性氧的淸除能力时衡量植物抗逆性的重要指标之一[20]。本实验中,短时间的干旱胁迫SOD活性增加,说明萱草可以通过自身的防御系统来维持细胞正常代谢活动;但随着胁迫程度的加剧和胁迫时间的延长,超出了萱草叶片的耐受程度,SOD活性急速下降,这与肖钢等[21]对夏玉米的研究结果基本一致。在复水7 d后,各干旱处理水平均恢复为对照水平,说明萱草具有较强的耐旱性。
叶绿素是植物进行光合作用的主要色素,其含量高低是表征光合速率的一个重要指标。实验中,叶绿素含量前期的升高可能由于水分胁迫造成萱草叶片含水量下降,叶片扩展生长受阻,致使叶绿素含量出现轻微上升,而干旱胁迫后期叶绿素含量的下降可能是因为持续干旱叶绿素的合成受到或者叶绿素分解。这与王宁等[22]对节节麦(AegilopstauschiiCoss)的研究结果一致。干旱胁迫解除后供水,轻度和重度干旱处理组均能恢复到正常水平,而重度干旱还是显著高于对照。这说明萱草叶片叶绿素的合成并未受到根本性破坏,在复水后可以通过自我调节机制进行恢复。
5 讨论
萱草在生理方面均对干旱胁迫和复水表现出显著响应,可以通过提高抗氧化酶活性和增加渗透调节物质含量来适应土壤水分亏缺,从而表现一定的耐旱能力。复水后,轻度和中度胁迫下萱草各指标都能恢复正常水平,说明旱后及时复水能缓解干旱胁迫对萱草生长发育的负面效应,具有一定的补偿效应,但其补偿强度不足以弥补重度干旱胁迫下萱草的生长发育。
猜你喜欢
科教新报(2022年24期)2022-07-08
大众健康(2021年7期)2021-07-28
安徽化工(2021年3期)2021-05-29
食品与生活(2020年8期)2020-09-12
科学导报(2020年39期)2020-06-21
现代农业科技(2020年10期)2020-06-04
科学与财富(2019年15期)2019-10-21
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
山东农业科学(2014年3期)2014-08-20
